
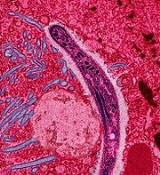
Plasmodium
Encyclopedia
Plasmodium is a genus of parasitic protist
s. Infection by these organisms is known as malaria
. The genus
Plasmodium was described in 1885 by Ettore Marchiafava
and Angelo Celli. Currently over 200 species of this genus are recognized and new species continue to be described.
Of the over 200 known species of Plasmodium, at least 11 species infect humans. Other species infect other animals, including monkey
s, rodent
s, bird
s, and reptile
s. The parasite always has two hosts in its life cycle
: a mosquito
vector and a vertebrate
host.
on November 6, 1880 at a military hospital in Constantine, Algeria
, when he discovered a microgametocyte exflagellating. In 1885, similar organisms were discovered within the blood of birds in Russia. There was brief speculation that birds might be involved in the transmission of malaria; in 1894 Patrick Manson
hypothesized that mosquito
es could transmit malaria. This hypothesis was independently confirmed by the Italian physician Giovanni Battista Grassi
working in Italy and the British physician Ronald Ross
working in India, both in 1898. Ross demonstrated the existence of Plasmodium in the wall of the midgut
and salivary glands of a Culex
mosquito using bird species as the vertebrate host. For this discovery he won the Nobel Prize
in 1902. Grassi showed that human malaria could only be transmitted by Anopheles
mosquitoes. It is worth noting, however, that for some species the vector may not be a mosquito.
, Plasmodium knowlesi
, Plasmodium vivax
and Plasmodium yoelii
- have been sequenced. All these species have genomes of about 25 megabases organised into 14 chromosomes consistent with earlier estimates. The chromosome
s vary in length from 500 kilobases to 3.5 megabases and it is presumed that this is the pattern throughout the genus.
The biology of these organisms is more fully described on the Plasmodium falciparum biology page
.
All the Plasmodium species causing malaria in humans are transmitted by mosquito species of the genus Anopheles. Species of the mosquito genera Aedes
, Culex, Culiseta
, Mansonia and Theobaldia can also transmit malaria but not to humans. Bird malaria is commonly carried by species belonging to the genus Culex. The life cycle of Plasmodium was discovered by Ross who worked with species from the genus Culex.
Both sexes of mosquitos live on nectar. Because nectar's protein content alone is insufficient for oogenesis (egg production) one or more blood meals is needed by the female. Only female mosquitoes bite.
Sporozoites from the saliva of a biting female mosquito are transmitted to either the blood or the lymphatic system of the recipient. It has been known for some time now that the parasites block the salivary ducts of the mosquito and as a consequence the insect normally requires multiple attempts to obtain blood. The reason for this has not been clear. It is now known that the multiple attempts by the mosquito may contribute to immunological tolerance of the parasite. The majority of sporozoites appear to be injected into the subcutaneous tissue from which they migrate into the capillaries. A proportion are ingested by macrophages and still others are taken up by the lymphatic system where they are presumably destroyed. ~10% of the parasites inoculated by the mosquitoes may remain in the skin where they may develop into infective merozoites.
and invade hepatocytes. For reasons that are currently unclear each sporozoite typically penetrates several hepatocytes before choosing one to reside within. Once the sporozoite has ceased migration it undergoes an initial remodelling of the pellicle, with disassembly of the inner membrane complex and the appearance of a bulb that progressively enlarges until the initially elongated sporozoite has transformed into a rounded form. This rounded form then matures within the hepatocyte to a schizont containing many merozoites. In some Plasmodium species, such as Plasmodium vivax and Plasmodium ovale
, the parasite in the hepatocyte may not achieve maturation to a schizont immediately but remain as a latent or dormant form and called a hypnozoite. Although Plasmodium falciparum is not considered to have a hypnozoite form, this may not be entirely correct (vide infra). This stage may be as short as 48 hours in the rodent parasites and as long as 15 days in P. malariae in humans.
There is considerable variation in the appearance of the blood forms between individuals experimentally inoculated at the same time. Even within a single experimentally individual there may be considerable variation in the maturity of the hepatic forms seen on liver biopsy.
A proportion of the hepatic stages may remain within the liver for considerable time - a form known as hypnozoites. Reactivation of the hypnozoites has been reported for up to 30 years after the initial infection in humans. The factors precipating this reactivation are not known. In the species Plasmodium ovale and Plasmodium vivax, but not in Plasmodium malariae
, hypnozoites have been shown to occur. It is not yet known if hypnozoite reactivaction occurs with any of the remaining species that infect humans but this is presumed to be the case.
The development from the hepatic stages to the erythrocytic stages has, until very recently, been obscure. In 2006 it was shown that the parasite buds off the hepatocytes in merosomes containing hundreds or thousands of merozoites. These merosomes lodge in the pulmonary capillaries and slowly disintegrate there over 48–72 hours releasing merozoites. The membrane of the merosome is derived from the host hepatocyte. The membrane of the merozoites is formed by repeated invagination of the parasite's membrane. The parastitophorus vacuole breaks down within the hepatocyte. This is associated with degeneration of the host cell's mitochondria and cessation of protein synthesis which is probably due to the lack of mitochondially produced ATP. The membrane of the merosome is then formed from that of the hepatocyte membrane but the hepatocyte proteins within the membrane are lost. This host derived membrane presumably provides protection from the immune system while the merozoites are transported to the lung. Erythrocyte invasion is enhanced when blood flow is slow and the cells are tightly packed: both of these conditions are found in the alveolar capillaries.
Infection of the liver may be influenced by the iron regulatory hormone hepcidin
and this may play a role in preventing superinfection despite repeated inoculation.
becomes prominent. The nucleus may become lobulated.
Within the erythrocytes the merozoite grow first to a ring-shaped form and then to a larger trophozoite form. In the schizont stage, the parasite divides several times to produce new merozoites, which leave the red blood cells and travel within the bloodstream to invade new red blood cells. The parasite feeds by ingesting haemoglobin and other materials from red blood cells and serum. The feeding process damages the erythrocytes. Details of this process have not been studied in species other than Plasmodium falciparum so generalizations may be premature at this time.
Erythrocytes infected by Plasmodium falciparum tend to form clumps - rosettes - and these have been linked to pathology caused by vascular occlusion. This rosette formation may be inhibted by heparin
. This agent has been used in the past as part of the treatment of malaria but was abandoned because of an increased risk of haemorrhage. Low molecular weight heparin also disrupts rosette formation and may have a lower risk of bleeding in malaria.
gives rise to the apical rings and conoid
. As development proceeds an aggregation of smooth membranes and the nucleus enter the base of the bud. The cytoplasm
contains numerous large ribosomes. Synchronous multiple cytoplasmic cleavage of the mature schizont results in the formation of numerous uninucleate merozoites.
Escape of the merozoites from the erythrocyte has also been studied. The erythrocyte swells under osmotic pressure. A pore opens in the erythrocte membrane and 1-2 meorozites escape. This is followed by an eversion the entire erythrocyte membrane, an action that propels the merozoites into the blood stream.
Invasion of erythrocyte precursors has only recently been studied. The earliest stage susceptible to infection were the orthoblasts - the stage immediately preceding the reticulocyte
stage which in turn is the immediate precursor to the mature erythrocyte. Invasion of the erythrocyte is inhibited by angiotensin
2 Angiotensin 2 is normally metabolized by erythrocytes to angiotensin (Ang) IV and Ang-(1-7). Parasite infection decreased the Ang-(1-7) levels and completely abolished Ang IV formation. Ang-(1-7), like its parent molecule, is capable of decreasing the level of infection. The mechanism of inhibition seems likely to be an inhibition of protein kinase A activity within the erythrocyte.
s) (also in the blood), which are taken up by the female mosquito. This process of differentiation into gametocytes appears to occur in the bone marrow. Five distinct morphological stages have recognised (stages I - V). Female gametocytes are produced about four times as commonly as male. In chonic infections in humans the gametocytes are often the only forms found in the blood. Incidentally the characteristic form of the female gametocytes in Plasmodium falciparum gave rise to this species's name.
Gameteocytes appear in the blood after a number of days post infection. In P. falciparum infections they appear after 7 to 15 days while in others they appear after 1 to 3 days. The ratio of asexual to sexual forms is between 10:1 and 156:1 The half life of the gametocytes has been estimated to be between 2 and 3 days but some are known to persist for up to four weeks.
Gametocyte carriage is associated with anaemia. Although female gametocytes normally outnumber males this may be reversed in the presence of anaemia.
One constant feature of the gametocytes in all stages that distinguishes them from the asexual forms is the presence of a pellicular complex. This originates in small membranous vesicle observed beneath the gametocyte plasmalemma in late stage I. Its function is not known. The structure itself consists of a subpellicular membrane vacuole. Deep to this is an array of longitudinally oriented microtubules. This structure is likely to be relatively inflexible and may help to explain the lack of amoeboid forms observed in asexual parasites.
Early stage one gametoctyes are very difficult to distinguish from small round trophozoites. Later stages can be distinguished by the distribution of pigment granulues. Under the electrom microscope the formation of the subpellicular membrane and a smooth plasma membrane are recognisable. The nuclei are recognisably dimorphic into male and female. These forms may be found between day 0 and day 2 in P falciparum infections.
In stage two the gametocyte enlarges and becomes D shaped. The nucleus may occupy a terminal end of the cell or lie along its length. Early spindle formation may be visible. These forms are found between day 1 to day 4 in P falciparum infections.
In stage three the erythrocyte becomes distorted. A staining difference between the male and female gametoctyes is apparent (male stain pink while female stain faint blue with the usual stains). The male nucleus is noticeably larger than the female and more lobulated. The female cytoplasm has more ribosomes, endoplasmic reticulum and mitochondria.
In stage four the erythrocyte is clearly deformed and the gametocyte is elongated. The male gametocytes stain red while the female stain violet blue. In the male pigment granules are scattered while in the female they are more dense. In the male the kinetochores of each chromosome are located over a nuclear pore. Osmophilic bodies are found in both but are more numerous in the female. These forms are found between day 6 and day 10 in P falciparum infections.
In stage five the gametocytes are clearly recognisable on light microscopy with the typical banana shaped female gametocytes. The subpellicular microtubules depolymerise but the membrane itself remains. In the male gametocyte exhibit the is a dramatic reduction in ribosomal density. Very few mitochondria are retained and the nucleus enlarges with a kinetochore complex attached to the nuclear envelope. In the female gametocytes there are numerous mitochondria, ribosomes and osmophillic bodies. The nucleus is small with a transcription factory.
Stages other than stage five are not normally found in the periferal blood. For reasons not yet understood stages I to IV are sequestered preferentially in the bone marrow and spleen. Stage V gametocytes only become infectious to mosquitoes after a further two or three days of circulation.
s and fertilize each other, forming motile zygote
s called ookinetes. It has been shown that up to 50% of the ookinetes may undergo apoptosis within the midgut. The reason for this behavior is unknown. While in the mosquito gut the parasites form thin cytoplasmic extensions to communicate with each other. These structures persist from the time of gametocyte activation until the zygote transforms into an ookinete. The function of these tubular structres remains to be discovered.
The ookinetes penetrate and escape the midgut, then embed themselves onto the exterior of the gut membrane. As in the liver the parasite tends to invade a number of cells before choosing one to reside in. The reason for the behavior is not known. Here they divide many times to produce large numbers of tiny elongated sporozoites. These sporozoites migrate to the salivary glands of the mosquito where they are injected into the blood and subcutaneous tissue of the next host the mosquito bites.
The escape of the gametocytes from the erythrocytes has been until recently obscure. The parasitophorous vacuole membrane ruptures at multiple sites within less than a minute following ingestion. This process may be inhibited by cysteine
protease
inhibitors. After this rupture of the vacoule the subpellicular membrane begins to disintegrate. This process also can be inhibited by aspartic
and the cysteine/serine
protease inhibitors. Approximately 15 minutes post-activation, the erythrocyte membrane ruptures at a single breaking point a third process that can be interrupted by protease inhibitors.
Infection of the mosquito has noticeable effects on the host. The presence of the parasite induces apotosis of the egg follicles.
.
Under favourable conditions asexual reproduction is superior to sexual as the parent is well adapted to its environment and its descendents share these genes. Transferring to a new host or in times of stress, sexual reproduction is generally superior as this produces a shuffling of genes
which on average at a population level will produce individuals better adapted to the new environment.
The advantages to asexual reproduction within a host can be seen from this simple model taken from Cook. The proportion of hosts that are parasitised is assumed to be small. This being the case the Poisson distribution
is a reasonable model. If the parasite is self fertilizing then the chance of successful reproduction is 1 - e-m where m is the proportion of the population parasitised. If the parasite is a faculative bisexual one - one that requires the presence of another parasite on the same host the likelyhood of success is 1 - (1 + m)e-m. If the parasite has two distinct sexes and requires both for reproduction, then the chance of success is ∑ (1 - 21-n)(mn/n!e-m) where the sum is taken between n = 2 and infinity. If m = 0.1 then the chance of success of the self fertilizing parasite is 40 times that of one with distinct sexes. The chance of success of the bisexual parasite is twice that of the parasite with distinct sexes. For smaller values of m, the advantages of self fertilization are even greater.
Given that this parasite spends part of its life cycle in two different hosts it must use a proportion of its available resources within each host. The proportion utilized is currently unknown. Empiracal estimates of this parameter are desirable for modeling of its life cycle.
A third case of an apparent recurrence nine years after leaving an endemic area of P. falciparum malaria has now been reported. A fourth case of recurrence in a patient with lung cancer has been reported. Two cases in pregnant women both from Africa but who had not lived there for over a year have been reported.
A case of congenital malaria due to both P. falciparum and P. malariae has been reported in a child born to a woman from Ghana
, a malaria endemic area, despite the mother having emigrated to Austria
eighteen months before and never having returned. A second case of congenital malaria in twins due to P. falciparum has been reported. The mother had left Togo 14 months before the diagnosis, had not returned in the interim and was never diagnosed with malaria during her pregnancy.
One case of malaria has been reported in a man of African origin with sickle cell trait who was treated for B cell
lymphoma
with chemotherapy and an autologous bone marrow
transplant. He developed symptomatic malaria only after a subsequent splenectomy performed for worsening disease. Pre treatment blood films and antigen testing were negative.
It seems that at least occasionally P. falciparum has a dormant stage. If this is in fact the case, eradication or control of this organism may be more difficult than previously believed.
. The drug induces a subpopulation of ring stages into developmental arrest. At the molecular level this is associated with overexpression of heat shock and erythrocyte binding surface proteins with the reduced expression of a cell-cycle regulator and a DNA biosynthesis protein.
The schizont stage-infected erythrocyte in an experimental culture of P. falciparum, F32 was suppressed to a low level with the use of atovaquone. The parasites resumed growth several days after the drug was removed from the culture.
- an organism that causes murine malaria. Similar merophores were found in the polymorph leukocytes and macrophages of other murine malaria parasite, P. yoelii nigeriensis
and P. chabaudi chabaudi
. All these species unlike P. falciparum are known to produce hyponozoites that may cause a relapse. The finding of Landau et al. on the presence of malaria parasites inside lymphatics suggest a mechanism for the recrudescence and chronicity of malaria infection.
s — a large group of photosynthetic protists. It is thought that the ancestors of the Apicomplexa were originally prey organisms that evolved the ability to invade the intestinal cells and subsequently lost their photosynthetic ability. Many of the species within the Apicomplexia still possess plastid
s (the organelle in which photosynthesis occurs in photosynthetic eukaryotes), and some that lack plastids nonetheless have evidence of plastid genes within their genomes. In the majority of such species, the plastids are not capable of photosynthesis
. Their function is not known, but there is suggestive evidence that they may be involved in reproduction.
Some extant dinoflagellates, however, can invade the bodies of jellyfish
and continue to photosynthesize, which is possible because jellyfish bodies are almost transparent. In host organisms with opaque
bodies, such an ability would most likely rapidly be lost. The 2008 description of a photosynthetic protist related to the Apicomplexia with a functional plastid supports this hypothesis.
A recently identified apicomplexan found in Australian corals - Chromera velia
- has retained a photosynthetic plastid.
An alternative to the ingestion hypothesis is that the ancestor of this group was a predator like Colpodella
that uses myzocytosis as its feeding mechanism instead invaded the prey cells instead of ingesting them.
and Haemoproteus evolved from one or more Leucocytozoon species. Parasites of the genus Leucocytozoan infect white blood cells (leukocytes) and liver and spleen
cells, and are transmitted by 'black flies' (Simulium
species) — a large genus of flies related to the mosquitoes.
It is thought that Leucocytozoon evolved from a parasite that spread by the orofaecal route and which infected the intestinal wall. At some point this parasite evolved the ability to infect the liver. This pattern is seen in the genus Cryptosporidium
, to which Plasmodium is distantly related. At some later point this ancestor developed the ability to infect blood cells and to survive and infect mosquitoes. Once vector transmission was firmly established, the previous orofecal route of transmission was lost.
The pattern of orofaecal transmission with coincidental infection of the erythrocytes is seen in the genus Schellackia
. Species in this genus infect lizards. The usual route of transmission is orofaecal but the parasites can also infect erythrocytes if they traverse the intestinal wall. The infected erythrocytes may be ingested by mites. These infected mites may subsequently be eaten by other uninfected lizards whereupon the parasites emerge and infect these new hosts. Unlike Plasmodium no development occurs in the mite.
Molecular evidence suggests that a reptile - specifically a squamate - was the first vertebrate host of Plasmodium. Birds were the second vertebrate hosts with mammals being the most recent group of vertebrates infected.
Leukocytes, hepatocyte
s and most spleen cells actively phagocytose particulate matter, which makes the parasite's entry into the cell easier. The mechanism of entry of Plasmodium species into erythrocytes is still very unclear, as it takes place in less than 30 seconds. It is not yet known if this mechanism evolved before mosquitoes became the main vectors for transmission of Plasmodium.
The genus Plasmodium evolved (presumably from its Leucocytozoon ancestor) about 130 million years ago, a period that is coincidental with the rapid spread of the angiosperms (flowering plants). This expansion in the angiosperms is thought to be due to at least one gene duplication
event. It seems probable that the increase in the number of flowers led to an increase in the number of mosquitoes and their contact with vertebrates.
about 230 million years ago. There are over 3500 species recognized, but to date their evolution has not been well worked out, so a number of gaps in our knowledge of the evolution of Plasmodium remain.
There is evidence of a recent expansion of Anopheles gambiae
and Anopheles arabiensis populations in the late Pleistocene
in Nigeria
.
The reason why a relatively limited number of mosquitoes should be such successful vectors of multiple diseases is not yet known. It has been shown that, among the most common disease-spreading mosquitoes, the symbiont bacterium Wolbachia
are not normally present. It has been shown that infection with Wolbachia can reduce the ability of some viruses and Plasmodium to infect the mosquito, and that this effect is Wolbachia-strain specific.
Plasmodiidae (Levine, 1988), order
Haemosporidia and phylum
Apicomplexa. There are currently 450 recognised species
in this order. Many species of this order are undergoing reexamination of their taxonomy with DNA
analysis. It seems likely that many of these species will be re-assigned after these studies have been completed. For this reason the entire order is outlined here.
Order Haemosporida
Family Haemoproteidae
Family Garniidae
Family Leucocytozoidae
Family Plasmodiidae
are:
While this tree contains a considerable number of species, DNA sequences from many species in this genus have not been included - probably because they are not available yet. Because of this problem, this tree and any conclusions that can be drawn from it should be regarded as provisional.
Three additional trees are available from the American Museum of Natural History.
These trees agree with the Tree of Life. Because of their greater number of species in these trees, some additional inferences can be made:
It is also known that the species infecting humans do not form a single clade. In contrast, the species infecting Old World monkeys seem to form a clade. Plasmodium vivax may have originated in Asia
and the related species Plasmodium simium appears to be derived through a transfer from the human P. vivax to New World
monkey species in South America. This occurred during an indepth study of Howler Monkeys near São Paulo, Brasil.
Another tree concetrating on the species infecting the primates is available here: PLOS site
This tree shows that the 'African' (P. malaria and P. ovale) and 'Asian' (P.cynomogli, P. gonderi, P. semiovale and P. simium) species tend to cluster together into separate clades. P. vivax clusters with the 'Asian' species. The rodent species (P. bergei, P. chabaudi and P. yoelli) form a separate clade. As usual P. falciparum does not cluster with any other species. The bird species (P. juxtanucleare, P. gallinaceum and P. relictum) form a clade that is related to the included Leucocytozoon and Haemoproteus species.
A second tree can be found on the PLoS website: PLOS site This tree concentrates largely on the species infecting primates.
The three bird species included in this tree (P. gallinacium, P. juxtanucleare and P. relictum) form a clade.
Four species (P. billbrayi, P. billcollinsi, P. falciparum and P. reichenowi) form a clade within the subgenus Lavernia. This subgenus is more closely related to the other primate species than to the bird species or the included Leuocytozoan species. Both P. billbrayi and P. billcollinsi infect both the chimpanzee subspecies included in this study (Pan troglodytes troglodytes and Pan troglodytes schweinfurthii). P. falciparum infects the bonbo (Pan paniscus) and P. reichenowi infects only one subspecies (Pan troglodytes troglodytes).
The eleven 'Asian' species included here form a clade with P. simium and P. vivax being clearly closely related as are P. knowseli and P. coatneyi; similarly P. brazillium and P. malariae are related. P. hylobati and P. inui are closely related. P. fragile and P. gonderi appear to be more closely related to P. vivax than to P. malariae.
P. coatneyi and P. inui appear to be closely related to P. vivax.
P. ovale is more closely related to P. malariae than to P. vivax.
Within the 'Asian' clade are three unnamed potential species. One infects each of the two chimpanzee subspecies included in the study (Pan troglodytes troglodytes and Pan troglodytes schweinfurthii). These appear to be related to the P. vivax/P. simium clade.
Two unnamed potential species infect the bonbo (Pan paniscus) and these are related to the P. malariae/P. brazillium clade.
The avian species were discovered soon after the description of P. falciparum and a variety of generic names were created. These were subsequently placed into the genus Plasmodium although some workers continued to use the genera Laverinia and Proteosoma for P. falciparum and the avian species respectively. The 5th and 6th Congresses of Malaria held at Istanbul
(1953) and Lisbon
(1958) recommended the creation and use of subgenera in this genus. Laverinia was applied to the species infecting humans and Haemamoeba to those infecting lizards and birds. This proposal was not universally accepted. Bray in 1955 proposed a definition for the subgenus Plasmodium and a second for the subgenus Laverinia in 1958. Garnham described a third subgenus - Vinckeia - in 1964.
The type species is Plasmodium falciparum.
Species infecting monkeys and apes (the higher primates) other than those in the subgenus Laverania are placed in the subgenus Plasmodium. The position of the recently described Plasmodium GorA and Plasmodium GorB has not yet been settled. The distinction between P. falciparum and P. reichenowi and the other species infecting higher primates was based on the morphological findings but have since been confirmed by DNA analysis.
The type species is Plasmodium malariae.
Parasites infecting other mammal
s including lower primates (lemur
s and others) are classified in the subgenus Vinckeia. Vinckeia while previously considered to be something of a taxonomic 'rag bag' has been recently shown - perhaps rather surprisingly - to form a coherent grouping.
The type species is Plasmodium bubalis
.
The four subgenera Giovannolaia, Haemamoeba, Huffia and Novyella were created by Corradetti et al. for the known avian malarial species. A fifth—Bennettinia—was created in 1997 by Valkiunas. The relationships between the subgenera are the matter of current investigation. Martinsen et al. 's recent (2006) paper outlines what is currently (2007) known. The subgenera Haemamoeba, Huffia, and Bennettinia appear to be monphylitic. Novyella appears to be well defined with occasional exceptions. The subgenus Giovannolaia needs revision.
P. juxtanucleare is currently (2007) the only known member of the subgenus Bennettinia.
Nyssorhynchus is an extinct subgenus of Plasmodium. It has one known member - Plasmodium dominicum
In 1966 Garnham classified those with large schizonts as Sauramoeba, those with small schizonts as Carinamoeba and the single then known species infecting snakes (Plasmodium wenyoni
) as Ophidiella. He was aware of the arbitrariness of this system and that it might not prove to be biologically valid. Telford in 1988 used this scheme as the basis for the currently accepted (2007) system.
These species have since been divided in to 8 genera - Asiamoeba, Carinamoeba, Fallisia, Garnia
, Lacertamoeba, Ophidiella, Paraplasmodium and Sauramoeba. Three of these genera (Asiamoeba, Lacertamoeba and Paraplasmodium) were created by Telford in 1988. Another species (Billbraya australis) described in 1990 by Paperna and Landau and is the only known species in this genus. This species may turn out to be another subgenus of lizard infecting Plasmodium.
With the exception of P. elongatum the exoerythrocytic stages occur in the endothelial cells and those of the macrophage-lymphoid system. The exoerythrocytic stages of P. elongatum parasitise the blood forming cells.
The various subgenera are first distinguished on the basis of the morphology of the mature gametocytes. Those of subgenus Haemamoeba are round or oval while those of the subgenera Giovannolaia, Huffia and Novyella are elongated. These latter genera are distinguished on the basis of the size of the schizonts: Giovannolaia and Huffia have large schizonts while those of Novyella are small.
Species in the subgenus Bennettinia have the following characteristics:
The type species is Plasmodium juxtanucleare
.
Species in the subgenus Giovannolaia have the following characteristics:
The type species is Plasmodium circumflexum
.
Species in the subgenus Haemamoeba have the following characteristics:
The type species is Plasmodium relictum.
Species in the subgenus Huffia have the following characteristics:
The type species is Plasmodium elongatum.
Species in the subgenus Novyella have the following characteristics:
The type species is Plasmodium vaughani
.
Species in the subgenus Asiamoeba have the following characteristics:
Species in the subgenus Carinamoeba have the following characteristics:
The type species is Plasmodium minasense
.
Species in the subgenus Fallisia have the following characteristics:
Species in the subgenus Garnia have the following characteristics:
Species in the subgenus Lacertaemoba have the following characteristics:
Species in the subgenus Paraplasmodium have the following characteristics:
Species in the subgenus Sauramoeba have the following characteristics:
The type species is Plasmodium agamae
.
All species in Ophidiella infect snakes
The type species is Plasmodium weyoni.
Notes
Asiamoeba
Bennetinia
Carinamoeba
Giovannolaia
Haemamoeba
Huffia
Lacertamoeba
Laverania
Ophidiella
Novyella
Nyssorhynchus
Paraplasmodium
Plasmodium
Sauramoeba
Vinckeia
are rarely affected; a few species are known to infect bat
s, porcupine
s and squirrel
s; carnivore
s, insectivore
s and marsupial
s are not known to act as hosts.
The listing of host species among the reptiles has rarely been attempted. Ayala in 1978 listed 156 published accounts on 54 valid species and subspecies between 1909 and 1975. The regional breakdown was Africa: 30 reports on 9 species; Australia
, Asia & Oceania
: 12 reports on 6 species and 2 subspecies; Americas
: 116 reports on 37 species.
Because of the number of species parasited by Plasmodium further discussion has been broken down into following pages:
Other useful references
Protist
Protists are a diverse group of eukaryotic microorganisms. Historically, protists were treated as the kingdom Protista, which includes mostly unicellular organisms that do not fit into the other kingdoms, but this group is contested in modern taxonomy...
s. Infection by these organisms is known as malaria
Malaria
Malaria is a mosquito-borne infectious disease of humans and other animals caused by eukaryotic protists of the genus Plasmodium. The disease results from the multiplication of Plasmodium parasites within red blood cells, causing symptoms that typically include fever and headache, in severe cases...
. The genus
Genus
In biology, a genus is a low-level taxonomic rank used in the biological classification of living and fossil organisms, which is an example of definition by genus and differentia...
Plasmodium was described in 1885 by Ettore Marchiafava
Ettore Marchiafava
Ettore Marchiafava was an Italian physician and zoologist who worked on malariaEttore Marchiafava , was the personal doctor of three popes and the Royal House of Savoy, a senator and professor of Pathological Anatomy at the Sapienza University of Rome...
and Angelo Celli. Currently over 200 species of this genus are recognized and new species continue to be described.
Of the over 200 known species of Plasmodium, at least 11 species infect humans. Other species infect other animals, including monkey
Monkey
A monkey is a primate, either an Old World monkey or a New World monkey. There are about 260 known living species of monkey. Many are arboreal, although there are species that live primarily on the ground, such as baboons. Monkeys are generally considered to be intelligent. Unlike apes, monkeys...
s, rodent
Rodent
Rodentia is an order of mammals also known as rodents, characterised by two continuously growing incisors in the upper and lower jaws which must be kept short by gnawing....
s, bird
Bird
Birds are feathered, winged, bipedal, endothermic , egg-laying, vertebrate animals. Around 10,000 living species and 188 families makes them the most speciose class of tetrapod vertebrates. They inhabit ecosystems across the globe, from the Arctic to the Antarctic. Extant birds range in size from...
s, and reptile
Reptile
Reptiles are members of a class of air-breathing, ectothermic vertebrates which are characterized by laying shelled eggs , and having skin covered in scales and/or scutes. They are tetrapods, either having four limbs or being descended from four-limbed ancestors...
s. The parasite always has two hosts in its life cycle
Biological life cycle
A life cycle is a period involving all different generations of a species succeeding each other through means of reproduction, whether through asexual reproduction or sexual reproduction...
: a mosquito
Mosquito
Mosquitoes are members of a family of nematocerid flies: the Culicidae . The word Mosquito is from the Spanish and Portuguese for little fly...
vector and a vertebrate
Vertebrate
Vertebrates are animals that are members of the subphylum Vertebrata . Vertebrates are the largest group of chordates, with currently about 58,000 species described. Vertebrates include the jawless fishes, bony fishes, sharks and rays, amphibians, reptiles, mammals, and birds...
host.
History
The organism itself was first seen by LaveranCharles Louis Alphonse Laveran
Charles Louis Alphonse Laveran was a French physician.In 1880, while working in the military hospital in Constantine, Algeria, he discovered that the cause of malaria is a protozoan, after observing the parasites in a blood smear taken from a patient who had just died of malaria.He also helped...
on November 6, 1880 at a military hospital in Constantine, Algeria
Constantine, Algeria
Constantine is the capital of Constantine Province in north-eastern Algeria. It was the capital of the same-named French département until 1962. Slightly inland, it is about 80 kilometres from the Mediterranean coast, on the banks of Rhumel river...
, when he discovered a microgametocyte exflagellating. In 1885, similar organisms were discovered within the blood of birds in Russia. There was brief speculation that birds might be involved in the transmission of malaria; in 1894 Patrick Manson
Patrick Manson
Sir Patrick Manson was a Scottish physician who made important discoveries in parasitology and was the founder of the tropical medicine field....
hypothesized that mosquito
Mosquito
Mosquitoes are members of a family of nematocerid flies: the Culicidae . The word Mosquito is from the Spanish and Portuguese for little fly...
es could transmit malaria. This hypothesis was independently confirmed by the Italian physician Giovanni Battista Grassi
Giovanni Battista Grassi
Giovanni Battista Grassi was an Italian zoologist, known for work demonstrating that mosquitos carry the malaria parasite Plasmodium in their digestive tract, on the embryological development of honey bees, on parasites, particularly the vine parasite phylloxera, migrations and metamorphosis in...
working in Italy and the British physician Ronald Ross
Ronald Ross
Sir Ronald Ross KCB FRS was a British doctor who received the Nobel Prize for Physiology or Medicine in 1902 for his work on malaria. He was the first Indian-born person to win a Nobel Prize...
working in India, both in 1898. Ross demonstrated the existence of Plasmodium in the wall of the midgut
Midgut
The midgut is the portion of the embryo from which most of the intestines develop. After it bends around the superior mesenteric artery, it is called the "midgut loop"...
and salivary glands of a Culex
Culex
Culex is a genus of mosquito, and is important in that several species serve as vectors of important diseases, such as West Nile virus, filariasis, Japanese encephalitis, St. Louis encephalitis and avian malaria....
mosquito using bird species as the vertebrate host. For this discovery he won the Nobel Prize
Nobel Prize
The Nobel Prizes are annual international awards bestowed by Scandinavian committees in recognition of cultural and scientific advances. The will of the Swedish chemist Alfred Nobel, the inventor of dynamite, established the prizes in 1895...
in 1902. Grassi showed that human malaria could only be transmitted by Anopheles
Anopheles
Anopheles is a genus of mosquito. There are approximately 460 recognized species: while over 100 can transmit human malaria, only 30–40 commonly transmit parasites of the genus Plasmodium, which cause malaria in humans in endemic areas...
mosquitoes. It is worth noting, however, that for some species the vector may not be a mosquito.
Biology
The genome of four Plasmodium species - Plasmodium falciparumPlasmodium falciparum
Plasmodium falciparum is a protozoan parasite, one of the species of Plasmodium that cause malaria in humans. It is transmitted by the female Anopheles mosquito. Malaria caused by this species is the most dangerous form of malaria, with the highest rates of complications and mortality...
, Plasmodium knowlesi
Plasmodium knowlesi
Plasmodium knowlesi is a primate malaria parasite commonly found in Southeast Asia. It causes malaria in long-tailed macaques , but it may also infect humans, either naturally or artificially....
, Plasmodium vivax
Plasmodium vivax
Plasmodium vivax is a protozoal parasite and a human pathogen. The most frequent and widely distributed cause of recurring malaria, P. vivax is one of the four species of malarial parasite that commonly infect humans. It is less virulent than Plasmodium falciparum, which is the deadliest of the...
and Plasmodium yoelii
Plasmodium yoelii
Plasmodium yoelii is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. yoelii has both vertebrate and insect hosts. The vertebrate hosts for this parasite are mammals.- Notes :...
- have been sequenced. All these species have genomes of about 25 megabases organised into 14 chromosomes consistent with earlier estimates. The chromosome
Chromosome
A chromosome is an organized structure of DNA and protein found in cells. It is a single piece of coiled DNA containing many genes, regulatory elements and other nucleotide sequences. Chromosomes also contain DNA-bound proteins, which serve to package the DNA and control its functions.Chromosomes...
s vary in length from 500 kilobases to 3.5 megabases and it is presumed that this is the pattern throughout the genus.
The biology of these organisms is more fully described on the Plasmodium falciparum biology page
Plasmodium falciparum biology
Plasmodium falciparum has been the focus of much research due to it being the causative agent of malaria. This article describes some of the recent findings surrounding the unique biology of this organism.-Life cycle:...
.
Diagnostic characteristics of the genus Plasmodium
- Merogony occurs both in erythrocytes and other tissues
- Merozoites, schizonts or gametocytes can be seen within erythrocytes and may displace the host nucleus
- Merozoites have a “signet-ring” appearance due to a large vacuole that forces the parasite’s nucleus to one pole
- Schizonts are round to oval inclusions that contain the deeply staining merozoites
- Forms gamonts in erythrocytes
- Gametocytes are 'halter-shaped' similar to HaemoproteusHaemoproteusHaemoproteus is a genus of protozoa that are parasitic in birds, reptiles and amphibians. Its name is derived from Greek: Haima - blood and Proteus - a sea god who had the power of assuming different shapes. The name Haemoproteus was first used in the description of Haemoproteus columbae in the...
but the pigment granules are more confined - Hemozoin is present
- Vectors are either mosquitos or sandflies
- Vertebrate hosts include mammals, birds and reptiles
Life cycle
The life cycle of Plasmodium while complex is similar to several other species in the Haemosporidia.All the Plasmodium species causing malaria in humans are transmitted by mosquito species of the genus Anopheles. Species of the mosquito genera Aedes
Aedes
Aedes is a genus of mosquito originally found in tropical and subtropical zones, but now found on all continents excluding Antarctica. Some species have been spread by human activity. Aedes albopictus, a most invasive species was recently spread to the New World, including the U.S., by the used...
, Culex, Culiseta
Culiseta
Culiseta is a genus of mosquitoes. Most Culiseta are cold-adapted species which only occur in warmer climates during the colder parts of the year or at higher elevations where temperatures are low...
, Mansonia and Theobaldia can also transmit malaria but not to humans. Bird malaria is commonly carried by species belonging to the genus Culex. The life cycle of Plasmodium was discovered by Ross who worked with species from the genus Culex.
Both sexes of mosquitos live on nectar. Because nectar's protein content alone is insufficient for oogenesis (egg production) one or more blood meals is needed by the female. Only female mosquitoes bite.
Sporozoites from the saliva of a biting female mosquito are transmitted to either the blood or the lymphatic system of the recipient. It has been known for some time now that the parasites block the salivary ducts of the mosquito and as a consequence the insect normally requires multiple attempts to obtain blood. The reason for this has not been clear. It is now known that the multiple attempts by the mosquito may contribute to immunological tolerance of the parasite. The majority of sporozoites appear to be injected into the subcutaneous tissue from which they migrate into the capillaries. A proportion are ingested by macrophages and still others are taken up by the lymphatic system where they are presumably destroyed. ~10% of the parasites inoculated by the mosquitoes may remain in the skin where they may develop into infective merozoites.
Hepatic stages
The majority of sporozoites migrate to the liverLiver
The liver is a vital organ present in vertebrates and some other animals. It has a wide range of functions, including detoxification, protein synthesis, and production of biochemicals necessary for digestion...
and invade hepatocytes. For reasons that are currently unclear each sporozoite typically penetrates several hepatocytes before choosing one to reside within. Once the sporozoite has ceased migration it undergoes an initial remodelling of the pellicle, with disassembly of the inner membrane complex and the appearance of a bulb that progressively enlarges until the initially elongated sporozoite has transformed into a rounded form. This rounded form then matures within the hepatocyte to a schizont containing many merozoites. In some Plasmodium species, such as Plasmodium vivax and Plasmodium ovale
Plasmodium ovale
Plasmodium ovale is a species of parasitic protozoa that causes tertian malaria in humans. It is closely related to Plasmodium falciparum and Plasmodium vivax, which are responsible for most malaria. It is rare compared to these two parasites, and substantially less dangerous than P...
, the parasite in the hepatocyte may not achieve maturation to a schizont immediately but remain as a latent or dormant form and called a hypnozoite. Although Plasmodium falciparum is not considered to have a hypnozoite form, this may not be entirely correct (vide infra). This stage may be as short as 48 hours in the rodent parasites and as long as 15 days in P. malariae in humans.
There is considerable variation in the appearance of the blood forms between individuals experimentally inoculated at the same time. Even within a single experimentally individual there may be considerable variation in the maturity of the hepatic forms seen on liver biopsy.
A proportion of the hepatic stages may remain within the liver for considerable time - a form known as hypnozoites. Reactivation of the hypnozoites has been reported for up to 30 years after the initial infection in humans. The factors precipating this reactivation are not known. In the species Plasmodium ovale and Plasmodium vivax, but not in Plasmodium malariae
Plasmodium malariae
Plasmodium malariae is a parasitic protozoa that causes malaria in humans. It is closely related to Plasmodium falciparum and Plasmodium vivax which are responsible for most malarial infection. While found worldwide, it is a so-called "benign malaria" and is not nearly as dangerous as that...
, hypnozoites have been shown to occur. It is not yet known if hypnozoite reactivaction occurs with any of the remaining species that infect humans but this is presumed to be the case.
The development from the hepatic stages to the erythrocytic stages has, until very recently, been obscure. In 2006 it was shown that the parasite buds off the hepatocytes in merosomes containing hundreds or thousands of merozoites. These merosomes lodge in the pulmonary capillaries and slowly disintegrate there over 48–72 hours releasing merozoites. The membrane of the merosome is derived from the host hepatocyte. The membrane of the merozoites is formed by repeated invagination of the parasite's membrane. The parastitophorus vacuole breaks down within the hepatocyte. This is associated with degeneration of the host cell's mitochondria and cessation of protein synthesis which is probably due to the lack of mitochondially produced ATP. The membrane of the merosome is then formed from that of the hepatocyte membrane but the hepatocyte proteins within the membrane are lost. This host derived membrane presumably provides protection from the immune system while the merozoites are transported to the lung. Erythrocyte invasion is enhanced when blood flow is slow and the cells are tightly packed: both of these conditions are found in the alveolar capillaries.
Infection of the liver may be influenced by the iron regulatory hormone hepcidin
Hepcidin
Hepcidin is a peptide hormone produced by the liver. It was discovered in 2000, and appears to be the master regulator of iron homeostasis in humans and other mammals. In humans, HAMP is the gene that encodes for hepcidin.-Structure:...
and this may play a role in preventing superinfection despite repeated inoculation.
Erythrocyte stages
After entering the erythrocyte, the merozoite lose one of their membranes, the apical rings, conoid and the rhopteries. Phagotropy commences and both smooth and granular endoplasmic reticulumEndoplasmic reticulum
The endoplasmic reticulum is an organelle of cells in eukaryotic organisms that forms an interconnected network of tubules, vesicles, and cisternae...
becomes prominent. The nucleus may become lobulated.
Within the erythrocytes the merozoite grow first to a ring-shaped form and then to a larger trophozoite form. In the schizont stage, the parasite divides several times to produce new merozoites, which leave the red blood cells and travel within the bloodstream to invade new red blood cells. The parasite feeds by ingesting haemoglobin and other materials from red blood cells and serum. The feeding process damages the erythrocytes. Details of this process have not been studied in species other than Plasmodium falciparum so generalizations may be premature at this time.
Erythrocytes infected by Plasmodium falciparum tend to form clumps - rosettes - and these have been linked to pathology caused by vascular occlusion. This rosette formation may be inhibted by heparin
Heparin
Heparin , also known as unfractionated heparin, a highly sulfated glycosaminoglycan, is widely used as an injectable anticoagulant, and has the highest negative charge density of any known biological molecule...
. This agent has been used in the past as part of the treatment of malaria but was abandoned because of an increased risk of haemorrhage. Low molecular weight heparin also disrupts rosette formation and may have a lower risk of bleeding in malaria.
Merozoites
The budding of the merozoites from interconnected cytoplasmic masses (pseudocytomeres) is a complex process. At the tip of each bud a thickened region of pelliclePellicle
Pellicle may refer to:*Pellicle , a thin layer supporting the cell membrane in various protozoa*Pellicle mirror, a thin plastic membrane which may be used as a beam splitter or protective cover in optical systems...
gives rise to the apical rings and conoid
Conoid
In geometry, a conoid is a Catalan surface all of whose rulings intersect a fixed line, called the axis of the conoid. If all its rulings are perpendicular to its axis, then the conoid is called a right conoid....
. As development proceeds an aggregation of smooth membranes and the nucleus enter the base of the bud. The cytoplasm
Cytoplasm
The cytoplasm is a small gel-like substance residing between the cell membrane holding all the cell's internal sub-structures , except for the nucleus. All the contents of the cells of prokaryote organisms are contained within the cytoplasm...
contains numerous large ribosomes. Synchronous multiple cytoplasmic cleavage of the mature schizont results in the formation of numerous uninucleate merozoites.
Escape of the merozoites from the erythrocyte has also been studied. The erythrocyte swells under osmotic pressure. A pore opens in the erythrocte membrane and 1-2 meorozites escape. This is followed by an eversion the entire erythrocyte membrane, an action that propels the merozoites into the blood stream.
Invasion of erythrocyte precursors has only recently been studied. The earliest stage susceptible to infection were the orthoblasts - the stage immediately preceding the reticulocyte
Reticulocyte
Reticulocytes are immature red blood cells, typically composing about 1% of the red cells in the human body.Reticulocytes develop and mature in the red bone marrow and then circulate for about a day in the blood stream before developing into mature red blood cells. Like mature red blood cells,...
stage which in turn is the immediate precursor to the mature erythrocyte. Invasion of the erythrocyte is inhibited by angiotensin
Angiotensin
Angiotensin, a peptide hormone, causes blood vessels to constrict, and drives blood pressure up. It is part of the renin-angiotensin system, which is a major target for drugs that lower blood pressure. Angiotensin also stimulates the release of aldosterone, another hormone, from the adrenal cortex...
2 Angiotensin 2 is normally metabolized by erythrocytes to angiotensin (Ang) IV and Ang-(1-7). Parasite infection decreased the Ang-(1-7) levels and completely abolished Ang IV formation. Ang-(1-7), like its parent molecule, is capable of decreasing the level of infection. The mechanism of inhibition seems likely to be an inhibition of protein kinase A activity within the erythrocyte.
Placental malaria
More than a hundred late-stage trophozoites or early schizont infected erythrocytes of P. falciparum in a case of placental malaria of a Tanzanian woman were found to form a nidus in an intervillous space of placenta. While such a concentration of parasites in placental malaria is rare, placental malaria cannot give rise to persistent infection as pregnancy in humans normally lasts only 9 months.Gametocytes
Most merozoites continue this replicative cycle but some merozoites differentiate into male or female sexual forms (gametocyteGametocyte
A gametocyte is a eukaryotic germ cell that divides by mitosis into other gametocytes or by meiosis into gametids during gametogenesis. Male gametocytes are called spermatocytes, and female gametocytes are called oocytes....
s) (also in the blood), which are taken up by the female mosquito. This process of differentiation into gametocytes appears to occur in the bone marrow. Five distinct morphological stages have recognised (stages I - V). Female gametocytes are produced about four times as commonly as male. In chonic infections in humans the gametocytes are often the only forms found in the blood. Incidentally the characteristic form of the female gametocytes in Plasmodium falciparum gave rise to this species's name.
Gameteocytes appear in the blood after a number of days post infection. In P. falciparum infections they appear after 7 to 15 days while in others they appear after 1 to 3 days. The ratio of asexual to sexual forms is between 10:1 and 156:1 The half life of the gametocytes has been estimated to be between 2 and 3 days but some are known to persist for up to four weeks.
Gametocyte carriage is associated with anaemia. Although female gametocytes normally outnumber males this may be reversed in the presence of anaemia.
Gametocyte morphology
The five recognised morphological stages were first described by Field and Shute in 1956.One constant feature of the gametocytes in all stages that distinguishes them from the asexual forms is the presence of a pellicular complex. This originates in small membranous vesicle observed beneath the gametocyte plasmalemma in late stage I. Its function is not known. The structure itself consists of a subpellicular membrane vacuole. Deep to this is an array of longitudinally oriented microtubules. This structure is likely to be relatively inflexible and may help to explain the lack of amoeboid forms observed in asexual parasites.
Early stage one gametoctyes are very difficult to distinguish from small round trophozoites. Later stages can be distinguished by the distribution of pigment granulues. Under the electrom microscope the formation of the subpellicular membrane and a smooth plasma membrane are recognisable. The nuclei are recognisably dimorphic into male and female. These forms may be found between day 0 and day 2 in P falciparum infections.
In stage two the gametocyte enlarges and becomes D shaped. The nucleus may occupy a terminal end of the cell or lie along its length. Early spindle formation may be visible. These forms are found between day 1 to day 4 in P falciparum infections.
In stage three the erythrocyte becomes distorted. A staining difference between the male and female gametoctyes is apparent (male stain pink while female stain faint blue with the usual stains). The male nucleus is noticeably larger than the female and more lobulated. The female cytoplasm has more ribosomes, endoplasmic reticulum and mitochondria.
In stage four the erythrocyte is clearly deformed and the gametocyte is elongated. The male gametocytes stain red while the female stain violet blue. In the male pigment granules are scattered while in the female they are more dense. In the male the kinetochores of each chromosome are located over a nuclear pore. Osmophilic bodies are found in both but are more numerous in the female. These forms are found between day 6 and day 10 in P falciparum infections.
In stage five the gametocytes are clearly recognisable on light microscopy with the typical banana shaped female gametocytes. The subpellicular microtubules depolymerise but the membrane itself remains. In the male gametocyte exhibit the is a dramatic reduction in ribosomal density. Very few mitochondria are retained and the nucleus enlarges with a kinetochore complex attached to the nuclear envelope. In the female gametocytes there are numerous mitochondria, ribosomes and osmophillic bodies. The nucleus is small with a transcription factory.
Stages other than stage five are not normally found in the periferal blood. For reasons not yet understood stages I to IV are sequestered preferentially in the bone marrow and spleen. Stage V gametocytes only become infectious to mosquitoes after a further two or three days of circulation.
Infection of mosquito
In the mosquito's midgut, the gametocytes develop into gameteGamete
A gamete is a cell that fuses with another cell during fertilization in organisms that reproduce sexually...
s and fertilize each other, forming motile zygote
Zygote
A zygote , or zygocyte, is the initial cell formed when two gamete cells are joined by means of sexual reproduction. In multicellular organisms, it is the earliest developmental stage of the embryo...
s called ookinetes. It has been shown that up to 50% of the ookinetes may undergo apoptosis within the midgut. The reason for this behavior is unknown. While in the mosquito gut the parasites form thin cytoplasmic extensions to communicate with each other. These structures persist from the time of gametocyte activation until the zygote transforms into an ookinete. The function of these tubular structres remains to be discovered.
The ookinetes penetrate and escape the midgut, then embed themselves onto the exterior of the gut membrane. As in the liver the parasite tends to invade a number of cells before choosing one to reside in. The reason for the behavior is not known. Here they divide many times to produce large numbers of tiny elongated sporozoites. These sporozoites migrate to the salivary glands of the mosquito where they are injected into the blood and subcutaneous tissue of the next host the mosquito bites.
The escape of the gametocytes from the erythrocytes has been until recently obscure. The parasitophorous vacuole membrane ruptures at multiple sites within less than a minute following ingestion. This process may be inhibited by cysteine
Cysteine
Cysteine is an α-amino acid with the chemical formula HO2CCHCH2SH. It is a non-essential amino acid, which means that it is biosynthesized in humans. Its codons are UGU and UGC. The side chain on cysteine is thiol, which is polar and thus cysteine is usually classified as a hydrophilic amino acid...
protease
Protease
A protease is any enzyme that conducts proteolysis, that is, begins protein catabolism by hydrolysis of the peptide bonds that link amino acids together in the polypeptide chain forming the protein....
inhibitors. After this rupture of the vacoule the subpellicular membrane begins to disintegrate. This process also can be inhibited by aspartic
Aspartic acid
Aspartic acid is an α-amino acid with the chemical formula HOOCCHCH2COOH. The carboxylate anion, salt, or ester of aspartic acid is known as aspartate. The L-isomer of aspartate is one of the 20 proteinogenic amino acids, i.e., the building blocks of proteins...
and the cysteine/serine
Serine
Serine is an amino acid with the formula HO2CCHCH2OH. It is one of the proteinogenic amino acids. By virtue of the hydroxyl group, serine is classified as a polar amino acid.-Occurrence and biosynthesis:...
protease inhibitors. Approximately 15 minutes post-activation, the erythrocyte membrane ruptures at a single breaking point a third process that can be interrupted by protease inhibitors.
Infection of the mosquito has noticeable effects on the host. The presence of the parasite induces apotosis of the egg follicles.
Discussion
The pattern of alternation of sexual and asexual reproduction which may seem confusing at first is a very common pattern in parasitic species. The evolutionary advantages of this type of life cycle were recognised by Gregor MendelGregor Mendel
Gregor Johann Mendel was an Austrian scientist and Augustinian friar who gained posthumous fame as the founder of the new science of genetics. Mendel demonstrated that the inheritance of certain traits in pea plants follows particular patterns, now referred to as the laws of Mendelian inheritance...
.
Under favourable conditions asexual reproduction is superior to sexual as the parent is well adapted to its environment and its descendents share these genes. Transferring to a new host or in times of stress, sexual reproduction is generally superior as this produces a shuffling of genes
Gênes
Gênes is the name of a département of the First French Empire in present Italy, named after the city of Genoa. It was formed in 1805, when Napoleon Bonaparte occupied the Republic of Genoa. Its capital was Genoa, and it was divided in the arrondissements of Genoa, Bobbio, Novi Ligure, Tortona and...
which on average at a population level will produce individuals better adapted to the new environment.
The advantages to asexual reproduction within a host can be seen from this simple model taken from Cook. The proportion of hosts that are parasitised is assumed to be small. This being the case the Poisson distribution
Poisson distribution
In probability theory and statistics, the Poisson distribution is a discrete probability distribution that expresses the probability of a given number of events occurring in a fixed interval of time and/or space if these events occur with a known average rate and independently of the time since...
is a reasonable model. If the parasite is self fertilizing then the chance of successful reproduction is 1 - e-m where m is the proportion of the population parasitised. If the parasite is a faculative bisexual one - one that requires the presence of another parasite on the same host the likelyhood of success is 1 - (1 + m)e-m. If the parasite has two distinct sexes and requires both for reproduction, then the chance of success is ∑ (1 - 21-n)(mn/n!e-m) where the sum is taken between n = 2 and infinity. If m = 0.1 then the chance of success of the self fertilizing parasite is 40 times that of one with distinct sexes. The chance of success of the bisexual parasite is twice that of the parasite with distinct sexes. For smaller values of m, the advantages of self fertilization are even greater.
Given that this parasite spends part of its life cycle in two different hosts it must use a proportion of its available resources within each host. The proportion utilized is currently unknown. Empiracal estimates of this parameter are desirable for modeling of its life cycle.
Plasmodium falciparum malaria
A report of P. falciparum malaria in a patient with sickle cell anemia four years after exposure to the parasite has been published. A second report that P. falciparum malaria had become symptomatic eight years after leaving an endemic area has also been published.A third case of an apparent recurrence nine years after leaving an endemic area of P. falciparum malaria has now been reported. A fourth case of recurrence in a patient with lung cancer has been reported. Two cases in pregnant women both from Africa but who had not lived there for over a year have been reported.
A case of congenital malaria due to both P. falciparum and P. malariae has been reported in a child born to a woman from Ghana
Ghana
Ghana , officially the Republic of Ghana, is a country located in West Africa. It is bordered by Côte d'Ivoire to the west, Burkina Faso to the north, Togo to the east, and the Gulf of Guinea to the south...
, a malaria endemic area, despite the mother having emigrated to Austria
Austria
Austria , officially the Republic of Austria , is a landlocked country of roughly 8.4 million people in Central Europe. It is bordered by the Czech Republic and Germany to the north, Slovakia and Hungary to the east, Slovenia and Italy to the south, and Switzerland and Liechtenstein to the...
eighteen months before and never having returned. A second case of congenital malaria in twins due to P. falciparum has been reported. The mother had left Togo 14 months before the diagnosis, had not returned in the interim and was never diagnosed with malaria during her pregnancy.
One case of malaria has been reported in a man of African origin with sickle cell trait who was treated for B cell
B cell
B cells are lymphocytes that play a large role in the humoral immune response . The principal functions of B cells are to make antibodies against antigens, perform the role of antigen-presenting cells and eventually develop into memory B cells after activation by antigen interaction...
lymphoma
Lymphoma
Lymphoma is a cancer in the lymphatic cells of the immune system. Typically, lymphomas present as a solid tumor of lymphoid cells. Treatment might involve chemotherapy and in some cases radiotherapy and/or bone marrow transplantation, and can be curable depending on the histology, type, and stage...
with chemotherapy and an autologous bone marrow
Bone marrow
Bone marrow is the flexible tissue found in the interior of bones. In humans, bone marrow in large bones produces new blood cells. On average, bone marrow constitutes 4% of the total body mass of humans; in adults weighing 65 kg , bone marrow accounts for approximately 2.6 kg...
transplant. He developed symptomatic malaria only after a subsequent splenectomy performed for worsening disease. Pre treatment blood films and antigen testing were negative.
It seems that at least occasionally P. falciparum has a dormant stage. If this is in fact the case, eradication or control of this organism may be more difficult than previously believed.
Drug induced
Developmental arrest was induced by in vitro culture of P. falciparum in the presence of sub lethal concentrations of artemisininArtemisinin
Artemisinin , also known as Qinghaosu , and its derivatives are a group of drugs that possess the most rapid action of all current drugs against falciparum malaria. Treatments containing an artemisinin derivative are now standard treatment worldwide for falciparum malaria...
. The drug induces a subpopulation of ring stages into developmental arrest. At the molecular level this is associated with overexpression of heat shock and erythrocyte binding surface proteins with the reduced expression of a cell-cycle regulator and a DNA biosynthesis protein.
The schizont stage-infected erythrocyte in an experimental culture of P. falciparum, F32 was suppressed to a low level with the use of atovaquone. The parasites resumed growth several days after the drug was removed from the culture.
Biological refuges
Macrophages containing merozoites dispersed in their cytoplasm, called 'merophores', were observed in P. vinckei petteriPlasmodium vinckei
Plasmodium vinckei is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. vinckei has both vertebrate and insect hosts. The vertebrate hosts for this parasite are rodents.- Description :...
- an organism that causes murine malaria. Similar merophores were found in the polymorph leukocytes and macrophages of other murine malaria parasite, P. yoelii nigeriensis
Plasmodium yoelii
Plasmodium yoelii is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. yoelii has both vertebrate and insect hosts. The vertebrate hosts for this parasite are mammals.- Notes :...
and P. chabaudi chabaudi
Plasmodium chabaudi
Plasmodium chabaudi is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. chabaudi has both vertebrate and insect hosts. The vertebrate hosts for this parasite are rodents.- Description :...
. All these species unlike P. falciparum are known to produce hyponozoites that may cause a relapse. The finding of Landau et al. on the presence of malaria parasites inside lymphatics suggest a mechanism for the recrudescence and chronicity of malaria infection.
Evolution
, DNA sequences are available from less than sixty species of Plasmodium and most of these are from species infecting either rodent or primate hosts. The evolutionary outline given here should be regarded as speculative, and subject to revision as more data becomes available.Apicomplexa
The Apicomplexa (the phylum to which Plasmodium belongs) are thought to have originated within the DinoflagellateDinoflagellate
The dinoflagellates are a large group of flagellate protists. Most are marine plankton, but they are common in fresh water habitats as well. Their populations are distributed depending on temperature, salinity, or depth...
s — a large group of photosynthetic protists. It is thought that the ancestors of the Apicomplexa were originally prey organisms that evolved the ability to invade the intestinal cells and subsequently lost their photosynthetic ability. Many of the species within the Apicomplexia still possess plastid
Plastid
Plastids are major organelles found in the cells of plants and algae. Plastids are the site of manufacture and storage of important chemical compounds used by the cell...
s (the organelle in which photosynthesis occurs in photosynthetic eukaryotes), and some that lack plastids nonetheless have evidence of plastid genes within their genomes. In the majority of such species, the plastids are not capable of photosynthesis
Photosynthesis
Photosynthesis is a chemical process that converts carbon dioxide into organic compounds, especially sugars, using the energy from sunlight. Photosynthesis occurs in plants, algae, and many species of bacteria, but not in archaea. Photosynthetic organisms are called photoautotrophs, since they can...
. Their function is not known, but there is suggestive evidence that they may be involved in reproduction.
Some extant dinoflagellates, however, can invade the bodies of jellyfish
Jellyfish
Jellyfish are free-swimming members of the phylum Cnidaria. Medusa is another word for jellyfish, and refers to any free-swimming jellyfish stages in the phylum Cnidaria...
and continue to photosynthesize, which is possible because jellyfish bodies are almost transparent. In host organisms with opaque
Opacity (optics)
Opacity is the measure of impenetrability to electromagnetic or other kinds of radiation, especially visible light. In radiative transfer, it describes the absorption and scattering of radiation in a medium, such as a plasma, dielectric, shielding material, glass, etc...
bodies, such an ability would most likely rapidly be lost. The 2008 description of a photosynthetic protist related to the Apicomplexia with a functional plastid supports this hypothesis.
A recently identified apicomplexan found in Australian corals - Chromera velia
Chromera velia
Chromera velia, also known as a "chromerid", is a photosynthetic alga in the superphylum Alveolata.It has typical features of alveolates, being phylogenetically related to apicomplexa, and contains a photosynthetic plastid. C. velia uses this organelle as the primary energy source, thus being the...
- has retained a photosynthetic plastid.
An alternative to the ingestion hypothesis is that the ancestor of this group was a predator like Colpodella
Colpodella
Colpodella is a genus of alveolates comprising 5 species, and two further possible species: They share all the synapomorphies of apicomplexans, but are free-living, rather than parasitic. This genus was previously known as Spiromonas...
that uses myzocytosis as its feeding mechanism instead invaded the prey cells instead of ingesting them.
Plasmodium genus
Current (2007) theory suggests that the genera Plasmodium, HepatocystisHepatocystis
Hepatocystis is a genus of parasitic protozoa transmitted by midges of the Culicoides genus. Hosts include Old World primates, bats, hippopotamus and squirrels. This genus is not found in the New World...
and Haemoproteus evolved from one or more Leucocytozoon species. Parasites of the genus Leucocytozoan infect white blood cells (leukocytes) and liver and spleen
Spleen
The spleen is an organ found in virtually all vertebrate animals with important roles in regard to red blood cells and the immune system. In humans, it is located in the left upper quadrant of the abdomen. It removes old red blood cells and holds a reserve of blood in case of hemorrhagic shock...
cells, and are transmitted by 'black flies' (Simulium
Simulium
Simulium is a genus of black flies, which may transmit diseases such as onchocerciasis . It is a large genus with several hundred species, and 41 sub-genera....
species) — a large genus of flies related to the mosquitoes.
It is thought that Leucocytozoon evolved from a parasite that spread by the orofaecal route and which infected the intestinal wall. At some point this parasite evolved the ability to infect the liver. This pattern is seen in the genus Cryptosporidium
Cryptosporidium
Cryptosporidium is a protozoan that can cause gastro-intestinal illness with diarrhea in humans.Cryptosporidium is the organism most commonly isolated in HIV positive patients presenting with diarrhea...
, to which Plasmodium is distantly related. At some later point this ancestor developed the ability to infect blood cells and to survive and infect mosquitoes. Once vector transmission was firmly established, the previous orofecal route of transmission was lost.
The pattern of orofaecal transmission with coincidental infection of the erythrocytes is seen in the genus Schellackia
Schellackia
Schellackia is a genus of parasitic protozoa within the phylum Apicomplexa. The genus was described in 1919 by Reichenow. Species in this genus infect reptiles and amphibians.The type species is Schellackia bolivari.-Life cycle:...
. Species in this genus infect lizards. The usual route of transmission is orofaecal but the parasites can also infect erythrocytes if they traverse the intestinal wall. The infected erythrocytes may be ingested by mites. These infected mites may subsequently be eaten by other uninfected lizards whereupon the parasites emerge and infect these new hosts. Unlike Plasmodium no development occurs in the mite.
Molecular evidence suggests that a reptile - specifically a squamate - was the first vertebrate host of Plasmodium. Birds were the second vertebrate hosts with mammals being the most recent group of vertebrates infected.
Leukocytes, hepatocyte
Hepatocyte
A hepatocyte is a cell of the main tissue of the liver. Hepatocytes make up 70-80% of the liver's cytoplasmic mass.These cells are involved in:* Protein synthesis* Protein storage* Transformation of carbohydrates...
s and most spleen cells actively phagocytose particulate matter, which makes the parasite's entry into the cell easier. The mechanism of entry of Plasmodium species into erythrocytes is still very unclear, as it takes place in less than 30 seconds. It is not yet known if this mechanism evolved before mosquitoes became the main vectors for transmission of Plasmodium.
The genus Plasmodium evolved (presumably from its Leucocytozoon ancestor) about 130 million years ago, a period that is coincidental with the rapid spread of the angiosperms (flowering plants). This expansion in the angiosperms is thought to be due to at least one gene duplication
Gene duplication
Gene duplication is any duplication of a region of DNA that contains a gene; it may occur as an error in homologous recombination, a retrotransposition event, or duplication of an entire chromosome.The second copy of the gene is often free from selective pressure — that is, mutations of it have no...
event. It seems probable that the increase in the number of flowers led to an increase in the number of mosquitoes and their contact with vertebrates.
Vectors
Mosquitoes evolved in what is now South AmericaSouth America
South America is a continent situated in the Western Hemisphere, mostly in the Southern Hemisphere, with a relatively small portion in the Northern Hemisphere. The continent is also considered a subcontinent of the Americas. It is bordered on the west by the Pacific Ocean and on the north and east...
about 230 million years ago. There are over 3500 species recognized, but to date their evolution has not been well worked out, so a number of gaps in our knowledge of the evolution of Plasmodium remain.
There is evidence of a recent expansion of Anopheles gambiae
Anopheles gambiae
Anopheles gambiae is a complex of at least seven morphologically distinguishable species of mosquitoes in the genus Anopheles. This complex was recognised in the 1960s and includes the most important vectors of malaria in sub-Saharan Africa and the most efficient malaria vectors known.This species...
and Anopheles arabiensis populations in the late Pleistocene
Pleistocene
The Pleistocene is the epoch from 2,588,000 to 11,700 years BP that spans the world's recent period of repeated glaciations. The name pleistocene is derived from the Greek and ....
in Nigeria
Nigeria
Nigeria , officially the Federal Republic of Nigeria, is a federal constitutional republic comprising 36 states and its Federal Capital Territory, Abuja. The country is located in West Africa and shares land borders with the Republic of Benin in the west, Chad and Cameroon in the east, and Niger in...
.
The reason why a relatively limited number of mosquitoes should be such successful vectors of multiple diseases is not yet known. It has been shown that, among the most common disease-spreading mosquitoes, the symbiont bacterium Wolbachia
Wolbachia
Wolbachia is a genus of bacteria which infects arthropod species, including a high proportion of insects , as well as some nematodes. It is one of the world's most common parasitic microbes and is possibly the most common reproductive parasite in the biosphere...
are not normally present. It has been shown that infection with Wolbachia can reduce the ability of some viruses and Plasmodium to infect the mosquito, and that this effect is Wolbachia-strain specific.
Taxonomy
Plasmodium belongs to the familyFamily
In human context, a family is a group of people affiliated by consanguinity, affinity, or co-residence. In most societies it is the principal institution for the socialization of children...
Plasmodiidae (Levine, 1988), order
Order (biology)
In scientific classification used in biology, the order is# a taxonomic rank used in the classification of organisms. Other well-known ranks are life, domain, kingdom, phylum, class, family, genus, and species, with order fitting in between class and family...
Haemosporidia and phylum
Phylum
In biology, a phylum The term was coined by Georges Cuvier from Greek φῦλον phylon, "race, stock," related to φυλή phyle, "tribe, clan." is a taxonomic rank below kingdom and above class. "Phylum" is equivalent to the botanical term division....
Apicomplexa. There are currently 450 recognised species
Species
In biology, a species is one of the basic units of biological classification and a taxonomic rank. A species is often defined as a group of organisms capable of interbreeding and producing fertile offspring. While in many cases this definition is adequate, more precise or differing measures are...
in this order. Many species of this order are undergoing reexamination of their taxonomy with DNA
DNA
Deoxyribonucleic acid is a nucleic acid that contains the genetic instructions used in the development and functioning of all known living organisms . The DNA segments that carry this genetic information are called genes, but other DNA sequences have structural purposes, or are involved in...
analysis. It seems likely that many of these species will be re-assigned after these studies have been completed. For this reason the entire order is outlined here.
Order Haemosporida
Family Haemoproteidae
Haemoproteidae
The Haemoproteidae are a family of parasitic protozoa in the phylum Apicomplexia.The species in this family produce pigment and do not have an asexual cycle in the blood.There are three genera in this family...
- Genus HaemocystidiumHaemocystidiumHaemocystidium is a genus of parasitic protozoa belonging to the phylum Apicomplexia.-History:The genus Haemocystidium was created to give a name to the haemoproteid of a gecko belonging to the genus Hemidactylus in Sri Lanka by Castellani and Willey in 1904...
Castellani and Willey 1904, emend. Telford 1996
- Genus HaemoproteusHaemoproteusHaemoproteus is a genus of protozoa that are parasitic in birds, reptiles and amphibians. Its name is derived from Greek: Haima - blood and Proteus - a sea god who had the power of assuming different shapes. The name Haemoproteus was first used in the description of Haemoproteus columbae in the...
- Subgenus Parahaemoproteus
- Subgenus HaemoproteusHaemoproteusHaemoproteus is a genus of protozoa that are parasitic in birds, reptiles and amphibians. Its name is derived from Greek: Haima - blood and Proteus - a sea god who had the power of assuming different shapes. The name Haemoproteus was first used in the description of Haemoproteus columbae in the...
Family Garniidae
Garniidae
Garniidae is a family of parasitic protozoa in the phylum Apicomplexia.Like many species in the Apicomplexia all species in this family have two hosts in their life cycle – one in a vertebrate and one invertebrate...
- Genus FallisiaFallisiaFallisia is a genus of the family PlasmodiidaeThe genus was created by Lainson, Landau and Shaw in 1974. A revision of the genus Plasmodium by Levine in 1985 considered this subgenus to be a synonom of Plasmodium. The description of a new species Plasmodium siamense by Telford in 1986 lead to a...
Lainson, Landau & Shaw 1974- Subgenus Fallisia
- Subgenus Plasmodioides
- Genus GarniaGarniaGarnia is a genus of parasitic protozoa belonging to the phylum Apicomplexia.-Description:Ulike members of the Plasmodiidae and Haemoproteidae, species in this genus do not produce pigment when developing in the erythrocytes....
- Genus ProgarniaProgarniaProgarnia is a genus of parasitic protozoa belonging to the phylum Apicomplexia.-Description:Merogony occurs principally in leucocytes and thrombocytes and to a lesser extent in the erythrocytes. No pigment is produced....
Family Leucocytozoidae
- Genus Leucocytozoon
- Subgenus Leucocytozoon
- Subgenus AkibaAkibaAkiba may refer to:*Akiba-kei, a Japanese slang term*Tadatoshi Akiba , mayor of Hiroshima* Akiba, a subgenus of protozoa in the genus LeucocytozoonIn entertainment:* Akiba , a 2006 Japanese film...
Family Plasmodiidae
Plasmodiidae
The Plasmodiidae are a family of apicomplexan parasites, including the type genus Plasmodium, which is responsible for malaria. This genus was created in 1903 by Mesnil.They are one of the four families in the order Haemosporida....
- Genus BioccalaBioccalaBioccala is a genus of parasitic protozoa belonging to the phylum Apicomplexia.- Host range :The only known host of these parasites is the intermediate roundleaf bat ....
- Genus BillbrayaBillbrayaBillbraya is a genus of parasitic protozoa. The genus lies within the phylum Apicomplexia.There is one recognised species in this genus: Billbraya australis.-Description:This genus was described in 1990 by Paperna and Landau...
- Genus DionisiaDionisiaDionisia is a genus of parasitic protozoa belonging to the phylum Apicomplexia.The type species of this genus is Dionisia bunoi.- Description :The genus was described by Landau et al. in 1980....
- Genus HepatocystisHepatocystisHepatocystis is a genus of parasitic protozoa transmitted by midges of the Culicoides genus. Hosts include Old World primates, bats, hippopotamus and squirrels. This genus is not found in the New World...
- Genus MesniliumMesniliumMesnilium is a genus of parasitic protozoa of belonging to the phylum Apicomplexia. Its vertebrate hosts are fish. The vectors are not presently known but are thought likely to be leeches.-Description:...
- Genus NycteriaNycteriaNycteria is a genus of parasitic protozoa of belonging to the phylum Apicomplexia. Its vertebrate hosts are bats. The vectors are not presently known.-Description:Schizonts are found in liver parenchymal cells...
- Genus Plasmodium
- Subgenus AsiamoebaAsiamoebaAsiamoeba is a subgenus of the genus Plasmodium - all of which are parastic protozoa. The subgenus was created by Telford in 1988. Species in this subgenus infect lizards.- Diagnostic features :...
Telford 1988 - Subgenus BennettiniaBennettiniaBennettinia is a subgenus of the genus Plasmodium - all of which are parastic protozoa. The subgenus was created in 1997 by Valkiunas. Species in this subgenus infect birds.- Diagnostic features :...
Valkiūnas 1997 - Subgenus CarinamoebaCarinamoebaCarinamoeba is a subgenus of the genus Plasmodium - all of which are parastic protozoa. The subgenus was created in 1966 by Garnham. Species in this subgenus infect reptiles.- Diagnostic features :...
Garnham 1966 - Subgenus GiovannolaiaGiovannolaiaGiovanolaia is a subgenus of the genus Plasmodium created by Corradetti et al. in 1963. The parasites within this subgenus infect birds.- Diagnostic features :Species in the subgenus Giovanolaia have the following characteristics:...
Corradetti, Garnham & Laird 1963 - Subgenus HaemamoebaHaemamoebaHaemamoeba is a subgenus of the genus Plasmodium - all of which are parastic protozoa. The subgenus was created in 1963 by created by Corradetti et al....
Grassi & Feletti 1890 - Subgenus HuffiaHuffiaHuffia is a subgenus of the genus Plasmodium - all of which are parasitic protozoa. The subgenus was created in 1963 by Corradetti et al....
Garnham & Laird 1963 - Subgenus LacertaemobaLacertaemobaLacertaemoba is a subgenus of the genus Plasmodium - all of which are parasitic protozoa. All species in this subgenus infect reptiles.This subgenus was created by Telford to refine the classification of species then given as Plasmodium tropiduri...
Telford 1988 - Subgenus LaveraniaLaveraniaLaverania is a subgenus of the genus Plasmodium. The subgenus was created in 1958 by Bray.The name was first proposed by Welch in 1897 as a genus name for the group now known as Plasmodium but for a variety of reasons the genus name Plasmodium was preferred.-History:The first non human primate...
Bray 1963 - Subgenus NovyellaNovyellaNovyella is a subgenus of the genus Plasmodium - all of which are parastic protozoa. The subgenus was created in 1963 by created by Corradetti et al.. Species in this subgenus infect birds...
Corradetti, Garnham & Laird 1963 - Subgenus OphidiellaOphidiellaOphidiella is a subgenus of the genus Plasmodium created in 1966 by Garnham. .It was created as a subgenus for the then only known species infecting snakes - Plasmodium wenyoni.- Species in this subgenus :Plasmodium pessoai...
Garnham 1966 - Subgenus Plasmodium Bray 1963 emend. Garnham 1964
- Subgenus ParaplasmodiumParaplasmodiumParaplasmodium is a subgenus of the genus Plasmodium - all of which are parastic protozoa. The subgenus was created by Telford in 1988. Species in this subgenus infect lizards.- Diagnostic features :...
Telford 1988 - Subgenus SauramoebaSauramoebaSauramoeba is a subgenus of the genus Plasmodium - all of which are parastic protozoa. The subgenus was created in 1966 by Garnham. Species in this subgenus infect reptiles.- Diagnostic features :...
Garnham 1966 - Subgenus VinckeiaVinckeiaVinckeia is a subgenus of the genus Plasmodium - all of which are parastic protozoa. The subgenus Vinckeia was created by Garnham in 1964 to accommodate the mammalian parasites other than those infecting the primates.- Diagnostic features :...
Garnham 1964
- Subgenus Asiamoeba
- Genus PolychromophilusPolychromophilusPolychromophilus is a genus of protozoa that infects bats. These parasite is transmitted by insect vectors most of whom are presently unknown...
- Genus RayellaRayellaRayella is a genus of parasitic protozoa of belonging to the phylum Apicomplexia. Its vertebrate hosts are flying squirrels. The vectors are not presently known .-Hosts:R. gigantica - spotted giant flying squirrel...
- Genus SaurocytozoonSaurocytozoonSaurocytozoon is a genus of parasitic protozoa. They infect reptiles and are thought to be transmitted by mosquitoes. Only two species have been described to date. Their relationship with the other members of this order is not presently clear....
Phylogenetic trees
The relationship between a number of these species can be seen on the Tree of Life website. Perhaps the most useful inferences that can be drawn from this phylogenetic treePhylogenetic tree
A phylogenetic tree or evolutionary tree is a branching diagram or "tree" showing the inferred evolutionary relationships among various biological species or other entities based upon similarities and differences in their physical and/or genetic characteristics...
are:
- P. falciparum and P. reichenowi (subgenus Laverania) branched off early in the evolution of this genus
- The genus Hepatocystis is nested within (paraphytic with) the genus Plasmodium
- The primate (subgenus Plasmodium) and rodent species (subgenus Vinckeia) form distinct groups
- The rodent and primate groups are relatively closely related
- The lizard and bird species are intermingled
- Although Plasmodium gallinaceum (subgenus Haemamoeba) and Plasmodium elongatum (subgenus Huffia) appear be related here there are so few bird species (three) included, this tree may not accurately reflect their real relationship.
- While no snake parasites have been included these are likely to group with the lizard-bird division
While this tree contains a considerable number of species, DNA sequences from many species in this genus have not been included - probably because they are not available yet. Because of this problem, this tree and any conclusions that can be drawn from it should be regarded as provisional.
Three additional trees are available from the American Museum of Natural History.
These trees agree with the Tree of Life. Because of their greater number of species in these trees, some additional inferences can be made:
- The genus Hepatocystis appears to lie within the primate-rodent clade
- The genus Haemoproteus appears lie within the bird-lizard clade
- The trees are consistent with the proposed origin of Plasmodium from Leucocytozoon
It is also known that the species infecting humans do not form a single clade. In contrast, the species infecting Old World monkeys seem to form a clade. Plasmodium vivax may have originated in Asia
Asia
Asia is the world's largest and most populous continent, located primarily in the eastern and northern hemispheres. It covers 8.7% of the Earth's total surface area and with approximately 3.879 billion people, it hosts 60% of the world's current human population...
and the related species Plasmodium simium appears to be derived through a transfer from the human P. vivax to New World
New World
The New World is one of the names used for the Western Hemisphere, specifically America and sometimes Oceania . The term originated in the late 15th century, when America had been recently discovered by European explorers, expanding the geographical horizon of the people of the European middle...
monkey species in South America. This occurred during an indepth study of Howler Monkeys near São Paulo, Brasil.
Another tree concetrating on the species infecting the primates is available here: PLOS site
This tree shows that the 'African' (P. malaria and P. ovale) and 'Asian' (P.cynomogli, P. gonderi, P. semiovale and P. simium) species tend to cluster together into separate clades. P. vivax clusters with the 'Asian' species. The rodent species (P. bergei, P. chabaudi and P. yoelli) form a separate clade. As usual P. falciparum does not cluster with any other species. The bird species (P. juxtanucleare, P. gallinaceum and P. relictum) form a clade that is related to the included Leucocytozoon and Haemoproteus species.
A second tree can be found on the PLoS website: PLOS site This tree concentrates largely on the species infecting primates.
The three bird species included in this tree (P. gallinacium, P. juxtanucleare and P. relictum) form a clade.
Four species (P. billbrayi, P. billcollinsi, P. falciparum and P. reichenowi) form a clade within the subgenus Lavernia. This subgenus is more closely related to the other primate species than to the bird species or the included Leuocytozoan species. Both P. billbrayi and P. billcollinsi infect both the chimpanzee subspecies included in this study (Pan troglodytes troglodytes and Pan troglodytes schweinfurthii). P. falciparum infects the bonbo (Pan paniscus) and P. reichenowi infects only one subspecies (Pan troglodytes troglodytes).
The eleven 'Asian' species included here form a clade with P. simium and P. vivax being clearly closely related as are P. knowseli and P. coatneyi; similarly P. brazillium and P. malariae are related. P. hylobati and P. inui are closely related. P. fragile and P. gonderi appear to be more closely related to P. vivax than to P. malariae.
P. coatneyi and P. inui appear to be closely related to P. vivax.
P. ovale is more closely related to P. malariae than to P. vivax.
Within the 'Asian' clade are three unnamed potential species. One infects each of the two chimpanzee subspecies included in the study (Pan troglodytes troglodytes and Pan troglodytes schweinfurthii). These appear to be related to the P. vivax/P. simium clade.
Two unnamed potential species infect the bonbo (Pan paniscus) and these are related to the P. malariae/P. brazillium clade.
Subgenera
The full taxonomic name of a species includes the subgenus but this is often omitted. The full name indicates some features of the morphology and type of host species. Sixteen subgenera are currently recognised.The avian species were discovered soon after the description of P. falciparum and a variety of generic names were created. These were subsequently placed into the genus Plasmodium although some workers continued to use the genera Laverinia and Proteosoma for P. falciparum and the avian species respectively. The 5th and 6th Congresses of Malaria held at Istanbul
Istanbul
Istanbul , historically known as Byzantium and Constantinople , is the largest city of Turkey. Istanbul metropolitan province had 13.26 million people living in it as of December, 2010, which is 18% of Turkey's population and the 3rd largest metropolitan area in Europe after London and...
(1953) and Lisbon
Lisbon
Lisbon is the capital city and largest city of Portugal with a population of 545,245 within its administrative limits on a land area of . The urban area of Lisbon extends beyond the administrative city limits with a population of 3 million on an area of , making it the 9th most populous urban...
(1958) recommended the creation and use of subgenera in this genus. Laverinia was applied to the species infecting humans and Haemamoeba to those infecting lizards and birds. This proposal was not universally accepted. Bray in 1955 proposed a definition for the subgenus Plasmodium and a second for the subgenus Laverinia in 1958. Garnham described a third subgenus - Vinckeia - in 1964.
Mammal infecting species
Two species in the subgenus Laverania are currently recognised: P. falciparum and P. reichenowi. Three additional species - Plasmodium billbrayi, Plasmodium billcollinsi and Plasmodium gaboni - may also exist (based on molecular data) but a full description of these species have not yet been published. The presence of elongated gametocytes in several of the avian subgenera and in Laverania in addition to a number of clinical features suggested that these might be closely related. This is no longer thought to be the case.The type species is Plasmodium falciparum.
Species infecting monkeys and apes (the higher primates) other than those in the subgenus Laverania are placed in the subgenus Plasmodium. The position of the recently described Plasmodium GorA and Plasmodium GorB has not yet been settled. The distinction between P. falciparum and P. reichenowi and the other species infecting higher primates was based on the morphological findings but have since been confirmed by DNA analysis.
The type species is Plasmodium malariae.
Parasites infecting other mammal
Mammal
Mammals are members of a class of air-breathing vertebrate animals characterised by the possession of endothermy, hair, three middle ear bones, and mammary glands functional in mothers with young...
s including lower primates (lemur
Lemur
Lemurs are a clade of strepsirrhine primates endemic to the island of Madagascar. They are named after the lemures of Roman mythology due to the ghostly vocalizations, reflective eyes, and the nocturnal habits of some species...
s and others) are classified in the subgenus Vinckeia. Vinckeia while previously considered to be something of a taxonomic 'rag bag' has been recently shown - perhaps rather surprisingly - to form a coherent grouping.
The type species is Plasmodium bubalis
Plasmodium bubalis
Plasmodium bubalis is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. bubalis has both vertebrate and insect hosts. The vertebrate hosts for this parasite are mammals.- Description :...
.
Bird infecting species
The remaining groupings are based on the morphology of the parasites. Revisions to this system are likely to occur in the future as more species are subject to analysis of their DNA.The four subgenera Giovannolaia, Haemamoeba, Huffia and Novyella were created by Corradetti et al. for the known avian malarial species. A fifth—Bennettinia—was created in 1997 by Valkiunas. The relationships between the subgenera are the matter of current investigation. Martinsen et al. 's recent (2006) paper outlines what is currently (2007) known. The subgenera Haemamoeba, Huffia, and Bennettinia appear to be monphylitic. Novyella appears to be well defined with occasional exceptions. The subgenus Giovannolaia needs revision.
P. juxtanucleare is currently (2007) the only known member of the subgenus Bennettinia.
Nyssorhynchus is an extinct subgenus of Plasmodium. It has one known member - Plasmodium dominicum
Reptile infecting species
Unlike the mammalian and bird malarias those species (more than 90 currently known) that infect reptiles have been more difficult to classify.In 1966 Garnham classified those with large schizonts as Sauramoeba, those with small schizonts as Carinamoeba and the single then known species infecting snakes (Plasmodium wenyoni
Plasmodium wenyoni
Plasmodium wenyoni is a parasite of the genus Plasmodium.Like all Plasmodium species P. wenyoni has both vertebrate and insect hosts. The vertebrate hosts for this parasite are reptiles.- Description :...
) as Ophidiella. He was aware of the arbitrariness of this system and that it might not prove to be biologically valid. Telford in 1988 used this scheme as the basis for the currently accepted (2007) system.
These species have since been divided in to 8 genera - Asiamoeba, Carinamoeba, Fallisia, Garnia
Garnia
Garnia is a genus of parasitic protozoa belonging to the phylum Apicomplexia.-Description:Ulike members of the Plasmodiidae and Haemoproteidae, species in this genus do not produce pigment when developing in the erythrocytes....
, Lacertamoeba, Ophidiella, Paraplasmodium and Sauramoeba. Three of these genera (Asiamoeba, Lacertamoeba and Paraplasmodium) were created by Telford in 1988. Another species (Billbraya australis) described in 1990 by Paperna and Landau and is the only known species in this genus. This species may turn out to be another subgenus of lizard infecting Plasmodium.
Bird infecting species
There are ~40 recognised bird species. Although over 50 species have been described, several have been rejected as being invalid.With the exception of P. elongatum the exoerythrocytic stages occur in the endothelial cells and those of the macrophage-lymphoid system. The exoerythrocytic stages of P. elongatum parasitise the blood forming cells.
The various subgenera are first distinguished on the basis of the morphology of the mature gametocytes. Those of subgenus Haemamoeba are round or oval while those of the subgenera Giovannolaia, Huffia and Novyella are elongated. These latter genera are distinguished on the basis of the size of the schizonts: Giovannolaia and Huffia have large schizonts while those of Novyella are small.
Species in the subgenus Bennettinia have the following characteristics:
The type species is Plasmodium juxtanucleare
Plasmodium juxtanucleare
Plasmodium juxtanucleare is a parasite of the genus Plasmodium subgenus Bennettinia.Like all Plasmodium species P. juxtanucleare has both vertebrate and insect hosts. The vertebrate hosts for this parasite are birds.-Description:...
.
Species in the subgenus Giovannolaia have the following characteristics:
- Schizonts contain plentiful cytoplasm, are larger than the host cell nucleus and frequently displace it. They are found only in mature erythrocytes.
- Gametocytes are elongated.
- Exoerythrocytic schizogony occurs in the mononuclear phagocyte system.
The type species is Plasmodium circumflexum
Plasmodium circumflexum
Plasmodium circumflexum is a parasite of the genus Plasmodium subgenus Giovannolaia.Like all Plasmodium species P. circumflexum has both vertebrate and insect hosts. The vertebrate hosts for this parasite are birds.-Description:...
.
Species in the subgenus Haemamoeba have the following characteristics:
- Mature schizonts are larger than the host cell nucleus and commonly displace it.
- Gametocytes are large, round, oval or irregular in shape and are substantially larger than the host nucleus.
The type species is Plasmodium relictum.
Species in the subgenus Huffia have the following characteristics:
- Mature schizonts, while varying in shape and size, contain plentiful cytoplasm and are commonly found in immature erthryocytes.
- Gametocytes are elongated.
The type species is Plasmodium elongatum.
Species in the subgenus Novyella have the following characteristics:
- Mature schizonts are either smaller than or only slightly larger than the host nucleus. They contain scanty cytoplasm.
- Gametocytes are elongated. Sexual stages in this subgenus resemble those of Haemoproteus.
- Exoerythrocytic schizogony occurs in the mononuclear phagocyte system
The type species is Plasmodium vaughani
Plasmodium vaughani
Plasmodium vaughani is a parasite of the genus Plasmodium subgenus Novyella.Like all Plasmodium species P. vaughani has both vertebrate and insect hosts. The vertebrate hosts for this parasite are birds.- Description :...
.
Reptile infecting species
All species in these subgenera infect lizards.Species in the subgenus Asiamoeba have the following characteristics:
Species in the subgenus Carinamoeba have the following characteristics:
- Schizonts normally give rise to less than 8 merozoites
- Schizonts are normally smaller than the host nucleus
The type species is Plasmodium minasense
Plasmodium minasense
Plasmodium minasense is a parasite of the genus Plasmodium subgenus Carinamoeba.Like all Plasmodium species P. minasense has both vertebrate and insect hosts. The vertebrate hosts for this parasite are lizards.- Description :...
.
Species in the subgenus Fallisia have the following characteristics:
- Non-pigmented asexual and gametocyte forms are found in leukocytes and thrombocytes
Species in the subgenus Garnia have the following characteristics:
- Pigment is absent
Species in the subgenus Lacertaemoba have the following characteristics:
Species in the subgenus Paraplasmodium have the following characteristics:
Species in the subgenus Sauramoeba have the following characteristics:
- Schizonts normally give rise to more than 8 merozoites
- Schizonts are normally larger than the host nucleus
- Non-pigmented gametocytes are typically the only forms found
- Pigmented forms may be found in the leukocytes occasionally
The type species is Plasmodium agamae
Plasmodium agamae
Plasmodium agamae is a parasite of the genus Plasmodium subgenus Sauramoeba.Like all Plasmodium species it has both vertebrate and insect hosts. The vertebrate host is the rainbow lizard Agama agama...
.
All species in Ophidiella infect snakes
The type species is Plasmodium weyoni.
Notes
- The erythrocytes of both reptiles and birds retain their nucleus, unlike those of mammals. The reason for the loss of the nucleus in mammalian erythocytes remains unknown.
Species listed by subgenera
The listing given here by subgenus is incomplete. A full listing of the species is avilable at Plasmodium species.Asiamoeba
- Plasmodium clelandiPlasmodium clelandiPlasmodium clelandi is a parasite of the genus Plasmodium subgenus Carinamoeba.Like all Plasmodium species P. clelandi has both vertebrate and insect hosts. The vertebrate hosts for this parasite are reptiles.- Description :...
- Plasmodium draconisPlasmodium draconisPlasmodium draconis is a parasite of the genus Plasmodium subgenus Asiamoeba.Like all Plasmodium species P. draconis has both vertebrate and insect hosts...
- Plasmodium lionatumPlasmodium lionatumPlasmodium lionatum is a parasite of the genus Plasmodium subgenus Asiamoeba.Like all Plasmodium species P. achiotense has both vertebrate and insect hosts. The vertebrate hosts for this parasite are lizards.- Description :...
- Plasmodium saurocordatum
- Plasmodium vastatorPlasmodium vastatorPlasmodium vastator is a parasite of the genus Plasmodium subgenus Asiamoeba.Like all Plasmodium species P. vastator has both vertebrate and insect hosts...
Bennetinia
- Plasmodium juxtanuclearePlasmodium juxtanuclearePlasmodium juxtanucleare is a parasite of the genus Plasmodium subgenus Bennettinia.Like all Plasmodium species P. juxtanucleare has both vertebrate and insect hosts. The vertebrate hosts for this parasite are birds.-Description:...
Carinamoeba
- Plasmodium basilisciPlasmodium basilisciPlasmodium basilisci is a parasite of the genus Plasmodium subgenus Carinamoeba.Like all Plasmodium species P. basilisci has both vertebrate and insect hosts. The vertebrate hosts for this parasite are reptiles.- Description :...
- Plasmodium clelandi
- Plasmodium lygosomaePlasmodium lygosomaePlasmodium lygosomae is a parasite of the genus Plasmodium subgenus Carinamoeba.Like all Plasmodium species P. lygosomae has both vertebrate and insect hosts...
- Plasmodium mabuiae
- Plasmodium minasense
- Plasmodium rhadinurumPlasmodium rhadinurumPlasmodium rhadinurum is a parasite of the genus Plasmodium subgenus Carinamoeba.Like all Plasmodium species P. rhadinurum has both vertebrate and insect hosts...
- Plasmodium volansPlasmodium volansPlasmodium volans is a parasite of the genus Plasmodium subgenus Carinamoeba.Like all Plasmodium species P. volans has both vertebrate and insect hosts...
Giovannolaia
- Plasmodium anasumPlasmodium anasumPlasmodium anasum is a species of the genus Plasmodium .Like all species in this genus it has both vertebrate and insect hosts. The vertebrate host are birds....
- Plasmodium circumflexum
- Plasmodium dissanaikeiPlasmodium dissanaikeiPlasmodium dissanaikei is a parasite of the genus Plasmodium subgenus Giovannolaia.Like all Plasmodium species P. dissanaikei has both vertebrate and insect hosts. The vertebrate hosts for this parasite are birds.- Description :...
- Plasmodium duraePlasmodium duraePlasmodium durae is a parasite of the genus Plasmodium subgenus Giovannolaia.Like all Plasmodium species P. durae has both vertebrate and insect hosts...
- Plasmodium fallaxPlasmodium fallaxPlasmodium fallax is a parasite of the genus Plasmodium subgenus Giovannolaia.Like all Plasmodium species P. fallax has both vertebrate and insect hosts...
- Plasmodium formosanum
- Plasmodium gabaldoniPlasmodium gabaldoniPlasmodium gabaldoni is a parasite of the genus Plasmodium subgenus Giovannolaia.Like all Plasmodium species P. gabaldoni has both vertebrate and insect hosts. The vertebrate hosts for this parasite are birds.- Description :...
- Plasmodium garnhamiPlasmodium garnhamiPlasmodium garnhami is a protozoan parasite of the genus Plasmodium.Like all Plasmodium species, P. garnhami has both vertebrate and insect hosts. The vertebrate hosts for this parasite are birds.- Clinical features and host pathology :...
- Plasmodium gundersiPlasmodium gundersiPlasmodium gundersi is a parasite of the genus Plasmodium subgenus Giovannolaia.Like all Plasmodium species P. gundersi has both vertebrate and insect hosts. The vertebrate hosts for this parasite are birds....
- Plasmodium hegneriPlasmodium hegneriPlasmodium hegneri is a parasite of the genus Plasmodium subgenus Giovannolaia.Like all Plasmodium species P. hegneri has both vertebrate and insect hosts...
- Plasmodium lophurae
- Plasmodium pedioecetii
- Plasmodium pinnotti
- Plasmodium polarePlasmodium polarePlasmodium polare is a parasite of the genus Plasmodium subgenus Giovannolaia.Like all Plasmodium species P. polare has both vertebrate and insect hosts...
Haemamoeba
- Plasmodium cathemerium
- Plasmodium coggeshalliPlasmodium coggeshalliPlasmodium coggeshalli is a parasite of the genus Plasmodium subgenus Haemamoeba.Like all Plasmodium species P. coggeshalli has both vertebrate and insect hosts. The vertebrate hosts for this parasite are birds....
- Plasmodium coturnixi
- Plasmodium elongatum
- Plasmodium gallinaceumPlasmodium gallinaceumPlasmodium gallinaceum is a species of the genus Plasmodium that causes malaria in poultry.- Description :...
- Plasmodium giovannolai
- Plasmodium lutziPlasmodium lutziPlasmodium lutzi is a parasite of the genus Plasmodium.Like all Plasmodium species P. lutzi has both vertebrate and insect hosts. The vertebrate hosts for this parasite are birds....
- Plasmodium matutinum
- Plasmodium paddaePlasmodium paddaePlasmodium paddae is a parasite of the genus Plasmodium subgenus Haemamoeba.Like all Plasmodium species P. paddae has both vertebrate and insect hosts. The vertebrate hosts for this parasite are birds....
- Plasmodium parvulum
- Plasmodium relictum
- Plasmodium tejera
Huffia
- Plasmodium elongatum
- Plasmodium hermaniPlasmodium hermaniPlasmodium hermani is a parasite of the genus Plasmodium subgenus Huffia.Like all Plasmodium species P. hermani has both vertebrate and insect hosts...
Lacertamoeba
- Plasmodium floridensePlasmodium floridensePlasmodium floridense is a parasite of the genus Plasmodium subgenus Lacertaemoba.Like all Plasmodium species P. floridense has both vertebrate and insect hosts...
- Plasmodium tropiduriPlasmodium tropiduriPlasmodium tropiduri is a parasite of the genus Plasmodium subgenus Lacertaemoba.Like all Plasmodium species P. tropiduri has both vertebrate and insect hosts...
Laverania
- Plasmodium billbrayi
- Plasmodium billcollinsi
- Plasmodium falciparumPlasmodium falciparumPlasmodium falciparum is a protozoan parasite, one of the species of Plasmodium that cause malaria in humans. It is transmitted by the female Anopheles mosquito. Malaria caused by this species is the most dangerous form of malaria, with the highest rates of complications and mortality...
- Plasmodium gaboni
- Plasmodium reichenowi
Ophidiella
- Plasmodium pessoaiPlasmodium pessoaiPlasmodium pessoai is a parasite of the genus Plasmodium.Like all Plasmodium species P. pessoai has both vertebrate and insect hosts. The vertebrate hosts for this parasite are reptiles.- Description :...
- Plasmodium tomodoniPlasmodium tomodoniPlasmodium tomodoni is a parasite of the genus Plasmodium.Like all Plasmodium species P. tomodoni has both vertebrate and insect hosts. The vertebrate hosts for this parasite are reptiles.- Clinical features and host pathology :...
- Plasmodium wenyoniPlasmodium wenyoniPlasmodium wenyoni is a parasite of the genus Plasmodium.Like all Plasmodium species P. wenyoni has both vertebrate and insect hosts. The vertebrate hosts for this parasite are reptiles.- Description :...
Novyella
- Plasmodium ashfordiPlasmodium ashfordiPlasmodium ashfordi is a species of the genus Plasmodium subgenus Novyella.Like all species in this genus it has both vertebrate and insect hosts...
- Plasmodium bertiiPlasmodium bertiiPlasmodium bertii is a parasite of the genus Plasmodium subgenus Novyella.Like all Plasmodium species P. bertii has both vertebrate and insect hosts. The vertebrate hosts for this parasite are birds.- Description :...
- Plasmodium bambusicolaiPlasmodium bambusicolaiPlasmodium bambusicolai is a species of the genus Plasmodium subgenus Novyella.Like all species in this genus it has both vertebrate and insect hosts. The vertebrate host are birds....
- Plasmodium columbae
- Plasmodium corradettiiPlasmodium corradettiiPlasmodium corradettii is a parasite of the genus Plasmodium subgenus Novyella.Like all Plasmodium species P. corradettii has both vertebrate and insect hosts. The vertebrate hosts for this parasite are birds....
- Plasmodium dissanaikeiPlasmodium dissanaikeiPlasmodium dissanaikei is a parasite of the genus Plasmodium subgenus Giovannolaia.Like all Plasmodium species P. dissanaikei has both vertebrate and insect hosts. The vertebrate hosts for this parasite are birds.- Description :...
- Plasmodium globularisPlasmodium globularisPlasmodium globularis is a parasite of the genus Plasmodium subgenus Novyella.Like all Plasmodium species P. globularis has both vertebrate and insect hosts. The vertebrate hosts for this parasite are birds....
- Plasmodium hexamerium
- Plasmodium jiangiPlasmodium jiangiPlasmodium jiangi is a parasite of the genus Plasmodium subgenus Novyella.Like all Plasmodium species P. jiangi has both vertebrate and insect hosts. The vertebrate hosts for this parasite are birds....
- Plasmodium kempi
- Plasmodium lucensPlasmodium lucensPlasmodium lucens is a parasite of the genus Plasmodium subgenus NovyellaLike all Plasmodium species P. lucens has both vertebrate and insect hosts. The vertebrate hosts for this parasite are birds....
- Plasmodium megaglobularisPlasmodium megaglobularisPlasmodium megaglobularis is a parasite of the genus Plasmodium subgenus Novyella.Like all Plasmodium species P. megaglobularis has both vertebrate and insect hosts. The vertebrate hosts for this parasite are birds....
- Plasmodium multivacuolarisPlasmodium multivacuolarisPlasmodium multivacuolaris is a parasite of the genus Plasmodium subgenus NovyellaLike all Plasmodium species P. multivacuolaris has both vertebrate and insect hosts. The vertebrate hosts for this parasite are birds....
- Plasmodium nucleophilum
- Plasmodium papernaiPlasmodium papernaiPlasmodium papernai is a parasite of the genus Plasmodium subgenus Novyella.Like all Plasmodium species P. papernai has both vertebrate and insect hosts. The vertebrate hosts for this parasite are birds....
- Plasmodium parahexameriumPlasmodium parahexameriumPlasmodium parahexamerium is a parasite of the genus Plasmodium subgenus Novyella.Like all Plasmodium species P. parahexamerium has both vertebrate and insect hosts. The vertebrate hosts for this parasite are birds....
- Plasmodium paranucleophilumPlasmodium paranucleophilumPlasmodium paranucleophilum is a parasite of the genus Plasmodium.Like all Plasmodium species P. paranucleophilum has both vertebrate and insect hosts. The vertebrate hosts for this parasite are birds....
- Plasmodium rouxi
- Plasmodium vaughaniPlasmodium vaughaniPlasmodium vaughani is a parasite of the genus Plasmodium subgenus Novyella.Like all Plasmodium species P. vaughani has both vertebrate and insect hosts. The vertebrate hosts for this parasite are birds.- Description :...
Nyssorhynchus
- Plasmodium dominicum
Paraplasmodium
- Plasmodium chiricahuaePlasmodium chiricahuaePlasmodium chiricahuae is a parasite of the genus Plasmodium subgenus Lacertaemoba.Like all Plasmodium species P. chiricahuae has both vertebrate and insect hosts...
- Plasmodium mexicanumPlasmodium mexicanumPlasmodium mexicanum is a parasite of the genus Plasmodium subgenus Paraplasmodium.Like all Plasmodium species P. mexicanum has both vertebrate and insect hosts...
- Plasmodium pifanoiPlasmodium pifanoiPlasmodium pifanoi is a parasite of the genus Plasmodium subgenus Paraplasmodium.Like all Plasmodium species P. pifanoi has both vertebrate and insect hosts. The vertebrate hosts for this parasite are lizards.- Description :...
Plasmodium
- Plasmodium bouillizePlasmodium bouillizePlasmodium bouillize is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. bouillize has both vertebrate and insect hosts. The vertebrate hosts for this parasite are mammals....
- Plasmodium brasilianum
- Plasmodium cercopitheciPlasmodium cercopitheciPlasmodium cercopitheci is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. cercopitheci has both vertebrate and insect hosts. The vertebrate hosts for this parasite are mammals....
- Plasmodium coatneyi
- Plasmodium cynomolgi
- Plasmodium eylesiPlasmodium eylesiPlasmodium eylesi is a parasite of the genus Plasmodium subgenus Plasmodium.Like all Plasmodium species P. eylesi has both vertebrate and insect hosts. The vertebrate hosts for this parasite are mammals.- Description :...
- Plasmodium fieldi
- Plasmodium fragile
- Plasmodium georgesiPlasmodium georgesiPlasmodium georgesi is a parasite of the genus Plasmodium subgenus Plasmodium.Like all Plasmodium species P. georgesi has both vertebrate and insect hosts...
- Plasmodium girardi
- Plasmodium gonderi
- Plasmodium gora
- Plasmodium gorb
- Plasmodium inui
- Plasmodium jefferyi
- Plasmodium joyeuxiPlasmodium joyeuxiPlasmodium joyeuxi is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. joyeuxi has both vertebrate and insect hosts. The vertebrate hosts for this parasite are mammals....
- Plasmodium knowlesiPlasmodium knowlesiPlasmodium knowlesi is a primate malaria parasite commonly found in Southeast Asia. It causes malaria in long-tailed macaques , but it may also infect humans, either naturally or artificially....
- Plasmodium hyobati
- Plasmodium malariae
- Plasmodium ovale
- Plasmodium petersi
- Plasmodium pitheci
- Plasmodium rhodiani
- Plasmodium schweitzi
- Plasmodium semiovale
- Plasmodium semnopitheciPlasmodium semnopitheciPlasmodium semnopitheci is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. semnopitheci has both vertebrate and insect hosts. The vertebrate hosts for this parasite are mammals....
- Plasmodium silvaticumPlasmodium silvaticumPlasmodium silvaticum is a parasite of the genus Plasmodium subgenus Plasmodium.Like all Plasmodium species P. silvaticum has both vertebrate and insect hosts...
- Plasmodium simium
- Plasmodium vivax
- Plasmodium youngiPlasmodium youngiPlasmodium youngi is a parasite of the genus Plasmodium subgenus Plasmodium.Like all Plasmodium species P. youngi has both vertebrate and insect hosts. The vertebrate hosts for this parasite are mammals....
Sauramoeba
- Plasmodium achiotensePlasmodium achiotensePlasmodium achiotense is a parasite of the genus Plasmodium subgenus Sauramoeba.Like all Plasmodium species P. achiotense has both vertebrate and insect hosts. The vertebrate hosts for this parasite are reptiles.- Description :...
- Plasmodium adunyinkai
- Plasmodium aeuminatumPlasmodium aeuminatumPlasmodium aeuminatum is a parasite of the genus Plasmodium subgenus Sauramoeba.Like all Plasmodium species it has both vertebrate and mosquito hosts. The vertebrate hosts for this species are reptiles....
- Plasmodium agamae
- Plasmodium balliPlasmodium balliPlasmodium balli is a parasite of the genus Plasmodium.Like all Plasmodium species it has vertebrate and insect hosts. The vertebrate hosts are lizards: the insect vector is not yet known.- Clinical features and host pathology :Host record:...
- Plasmodium beltraniPlasmodium beltraniPlasmodium beltrani is a parasite of the genus Plasmodium subgenus Sauramoeba.Like all Plasmodium species P. beltrani has both vertebrate and insect hosts...
- Plasmodium brumptiPlasmodium brumptiPlasmodium brumpti is a parasite of the genus Plasmodium subgenus Sauramoeba.Like all Plasmodium species P. brumpti has both vertebrate and insect hosts...
- Plasmodium cnemidophoriPlasmodium cnemidophoriPlasmodium cnemidophori is a parasite of the genus Plasmodium subgenus Sauramoeba.Like all Plasmodium species P. cnemidophori has both vertebrate and insect hosts...
- Plasmodium diploglossiPlasmodium diploglossiPlasmodium diploglossi is a parasite of the genus Plasmodium subgenus Sauramoeba.Like all Plasmodium species P. diploglossi has both vertebrate and insect hosts. The vertebrate hosts for this parasite are reptiles.- Description :...
- Plasmodium giganteumPlasmodium giganteumPlasmodium giganteum is a parasite of the genus Plasmodium subgenus Sauramoeba.Like all Plasmodium species P. giganteum has both vertebrate and insect hosts...
- Plasmodium heischiPlasmodium heischiPlasmodium heischi is a parasite of the genus Plasmodium subgenus Sauramoeba.Like all Plasmodium species P. heischi has both vertebrate and insect hosts...
- Plasmodium josephinaePlasmodium josephinaePlasmodium josephinae is a parasite of the genus Plasmodium subgenus Sauramoeba.Like all Plasmodium species P. josephinae has both vertebrate and insect hosts. The vertebrate hosts for this parasite are reptiles....
- Plasmodium pelaeziPlasmodium pelaeziPlasmodium pelaezi is a parasite of the genus Plasmodium subgenus Sauramoeba.Like all Plasmodium species P. pelaezi has both vertebrate and insect hosts...
- Plasmodium zonuriaePlasmodium zonuriaePlasmodium zonuriae is a parasite of the genus Plasmodium subgenus Sauramoeba.Like all Plasmodium species P. zonuriae has both vertebrate and insect hosts. The vertebrate hosts for this parasite are reptiles....
Vinckeia
- Plasmodium achromaticumPlasmodium achromaticumPlasmodium achromaticum is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. achromaticum has both vertebrate and insect hosts. The vertebrate hosts for this parasite are mammals....
- Plasmodium aegyptensisPlasmodium aegyptensisPlasmodium aegyptensis is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species it has vertebrate and insect hosts. The vertebrate host is the Egyptian grass rat . The insect vector for this species is not yet known....
- Plasmodium anomaluriPlasmodium anomaluriPlasmodium anomaluri is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species, P. anomaluri has both vertebrate and insect hosts. The vertebrate hosts for this parasite are mammals....
- Plasmodium atheruriPlasmodium atheruriPlasmodium atheruri is a species of the genus Plasmodium subgenus Vinckeia.Like all members of this genus it is parasitic on vertebrate and insect hosts...
- Plasmodium bergheiPlasmodium bergheiPlasmodium berghei is a unicellular parasite and it infects mammals other than humans.P. berghei is one of the four Plasmodium species that have been described in African murine rodents....
- Plasmodium booliatiPlasmodium booliatiPlasmodium booliati is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. booliati has both vertebrate and insect hosts. The vertebrate hosts for this parasite are mammals....
- Plasmodium brodeniPlasmodium brodeniPlasmodium brodeni is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. brodeni has both vertebrate and insect hosts. The vertebrate hosts for this parasite are mammals.-Description:...
- Plasmodium bubalis
- Plasmodium buckiPlasmodium buckiPlasmodium bucki is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. bucki has both vertebrate and insect hosts. The vertebrate hosts for this parasite are mammals.- Description :...
- Plasmodium capraePlasmodium capraePlasmodium caprae is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. caprae has both vertebrate and insect hosts. The vertebrate hosts for this parasite are domestic goats, Capra aegagrus. The specific epithet for P...
- Plasmodium cephalophiPlasmodium cephalophiPlasmodium cephalophi is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. cephalophi has both vertebrate and insect hosts...
- Plasmodium chabaudiPlasmodium chabaudiPlasmodium chabaudi is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. chabaudi has both vertebrate and insect hosts. The vertebrate hosts for this parasite are rodents.- Description :...
- Plasmodium coulangesiPlasmodium coulangesiPlasmodium coulangesi is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. coulangesi has both vertebrate and insect hosts...
- Plasmodium cyclopsiPlasmodium cyclopsiPlasmodium cyclopsi is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. cyclopsi has both vertebrate and insect hosts. The vertebrate hosts for this parasite are mammals....
- Plasmodium foleyiPlasmodium foleyiPlasmodium foleyi is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. foleyi has both vertebrate and insect hosts. The vertebrate hosts for this parasite are mammals.- Description :...
- Plasmodium girardi
- Plasmodium incertaePlasmodium incertaePlasmodium incertae is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. incertae has both vertebrate and insect hosts. The vertebrate hosts for this parasite are mammals....
- Plasmodium inopinatumPlasmodium inopinatumPlasmodium inopinatum is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. inopinatum has both vertebrate and insect hosts. The vertebrate hosts for this parasite are mammals....
- Plasmodium landauaePlasmodium landauaePlasmodium landauae is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. landauae has both vertebrate and insect hosts. The vertebrate hosts for this parasite are mammals....
- Plasmodium lemurisPlasmodium lemurisPlasmodium lemuris is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. lemuris has both vertebrate and insect hosts...
- Plasmodium melanipherumPlasmodium melanipherumPlasmodium melanipherum is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. melanipherum has both vertebrate and insect hosts. The vertebrate hosts for this parasite are mammals.- Description :...
- Plasmodium narayaniPlasmodium narayaniPlasmodium narayani is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. narayani has both vertebrate and insect hosts. The vertebrate hosts for this parasite are mammals.- Description :...
- Plasmodium odocoilei
- Plasmodium percygarnhamiPlasmodium percygarnhamiPlasmodium percygarnhami is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. percygarnhami has both vertebrate and insect hosts...
- Plasmodium pulmophiliumPlasmodium pulmophiliumPlasmodium pulmophilium is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. pulmophilium has both vertebrate and insect hosts. The vertebrate hosts for this parasite are mammals....
- Plasmodium sandoshamiPlasmodium sandoshamiPlasmodium achiotense is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. sandoshami has both vertebrate and insect hosts. The vertebrate hosts for this parasite are mammals.- Description :...
- Plasmodium traguliPlasmodium traguliPlasmodium traguli is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. traguli has both vertebrate and insect hosts...
- Plasmodium tyrioPlasmodium tyrioPlasmodium tyrio is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. tyrio has both vertebrate and insect hosts...
- Plasmodium uilenbergiPlasmodium uilenbergiPlasmodium uilenbergi is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. uilenbergi has both vertebrate and insect hosts...
- Plasmodium vinckeiPlasmodium vinckeiPlasmodium vinckei is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. vinckei has both vertebrate and insect hosts. The vertebrate hosts for this parasite are rodents.- Description :...
- Plasmodium watteniPlasmodium watteniPlasmodium watteni is a parasite of the genus Plasmodium subgenus Vinckeia.Like all Plasmodium species P. watteni has both vertebrate and insect hosts. The vertebrate hosts for this parasite are mammals....
- Plasmodium yoelli
Host range
Host range among the mammalian orders is non uniform. At least 29 species infect non human primates; rodents outside the tropical parts of AfricaAfrica
Africa is the world's second largest and second most populous continent, after Asia. At about 30.2 million km² including adjacent islands, it covers 6% of the Earth's total surface area and 20.4% of the total land area...
are rarely affected; a few species are known to infect bat
Bat
Bats are mammals of the order Chiroptera "hand" and pteron "wing") whose forelimbs form webbed wings, making them the only mammals naturally capable of true and sustained flight. By contrast, other mammals said to fly, such as flying squirrels, gliding possums, and colugos, glide rather than fly,...
s, porcupine
Porcupine
Porcupines are rodents with a coat of sharp spines, or quills, that defend or camouflage them from predators. They are indigenous to the Americas, southern Asia, and Africa. Porcupines are the third largest of the rodents, behind the capybara and the beaver. Most porcupines are about long, with...
s and squirrel
Squirrel
Squirrels belong to a large family of small or medium-sized rodents called the Sciuridae. The family includes tree squirrels, ground squirrels, chipmunks, marmots , flying squirrels, and prairie dogs. Squirrels are indigenous to the Americas, Eurasia, and Africa and have been introduced to Australia...
s; carnivore
Carnivore
A carnivore meaning 'meat eater' is an organism that derives its energy and nutrient requirements from a diet consisting mainly or exclusively of animal tissue, whether through predation or scavenging...
s, insectivore
Insectivore
An insectivore is a type of carnivore with a diet that consists chiefly of insects and similar small creatures. An alternate term is entomophage, which also refers to the human practice of eating insects....
s and marsupial
Marsupial
Marsupials are an infraclass of mammals, characterized by giving birth to relatively undeveloped young. Close to 70% of the 334 extant species occur in Australia, New Guinea, and nearby islands, with the remaining 100 found in the Americas, primarily in South America, but with thirteen in Central...
s are not known to act as hosts.
The listing of host species among the reptiles has rarely been attempted. Ayala in 1978 listed 156 published accounts on 54 valid species and subspecies between 1909 and 1975. The regional breakdown was Africa: 30 reports on 9 species; Australia
Australia
Australia , officially the Commonwealth of Australia, is a country in the Southern Hemisphere comprising the mainland of the Australian continent, the island of Tasmania, and numerous smaller islands in the Indian and Pacific Oceans. It is the world's sixth-largest country by total area...
, Asia & Oceania
Oceania
Oceania is a region centered on the islands of the tropical Pacific Ocean. Conceptions of what constitutes Oceania range from the coral atolls and volcanic islands of the South Pacific to the entire insular region between Asia and the Americas, including Australasia and the Malay Archipelago...
: 12 reports on 6 species and 2 subspecies; Americas
Americas
The Americas, or America , are lands in the Western hemisphere, also known as the New World. In English, the plural form the Americas is often used to refer to the landmasses of North America and South America with their associated islands and regions, while the singular form America is primarily...
: 116 reports on 37 species.
Because of the number of species parasited by Plasmodium further discussion has been broken down into following pages:
- Plasmodium species infecting humans and other primatesPlasmodium species infecting humans and other primates- Species of Plasmodium infecting humans : *Plasmodium falciparum *Plasmodium vivax...
- Plasmodium species infecting mammals other than primatesPlasmodium species infecting mammals other than primatesThe subgenus Vinckeia of Plasmodium was created by Garnham to accommodate the mammalian parasites other than those infecting primates. Species infecting lemurs have also been included in this subgenus....
- Plasmodium species infecting birdsPlasmodium species infecting birdsSpecies in five subgenera of Plasmodium infect birds - Bennettinia, Giovannolaia, Haemamoeba, Huffia and Novyella. Giovannolaia appears to be a polyphytic group and may be sudivided in the future.- Parasite listing :...
- Plasmodium species infecting reptilesPlasmodium species infecting reptilesOver 90 species and subspecies of Plasmodium infect lizards. They have been reported from over 3200 species of lizard but only 29 species of snake. Three species - P. pessoai, P. tomodoni and P. wenyoni - infect snakes...
Species reclassified into other genera
The following species have been assigned to the genus Plasmodium in the past:- Hepatocystis epomophoriHepatocystis epomophoriHepatocystis epomophori is a species of the genus Hepatocystis.Species in this genus are parasitic protozoa transmitted by flies of the genus Culicoides and infect mammals.-Description:...
- Hepatocystis kochi
- Hepatocystis limnotragi Van Denberghe 1937
- Hepatocystis pteropi Breinl 1911
- Hepatocystis ratufae Donavan 1920
- Hepatocystis vassali Laveran 1905
- Haemoemba praecox
- Haemoemba rousseleti
- Garnia gonatodi
- Fallisia siamense
Species of dubious validity
The following species are currently regarded as questionable validity (nomen dubium).- Plasmodium bitis
- Plasmodium bowiei
- Plasmodium brucei
- Plasmodium bufoni
- Plasmodium caprea
- Plasmodium carinii
- Plasmodium causi
- Plasmodium chalcidi
- Plasmodium chloropsidis
- Plasmodium centropi
- Plasmodium danilweskyi
- Plasmodium divergens
- Plasmodium effusum
- Plasmodium fabesia
- Plasmodium gambeli
- Plasmodium galinulae
- Plasmodium herodiadis
- Plasmodium limnotragi
- Plasmodium malariae raupachi
- Plasmodium metastaticum
- Plasmodium moruony
- Plasmodium periprocoti
- Plasmodium ploceii
- Plasmodium struthionis
Further reading
Standard reference books for the identification of Plasmodium species-
- This book is the standard reference work on malarial species classification even if it a little dated now. A number of additional species have been described since its publication.
Other useful references
External links
Some history of malaria - http://muse.jhu.edu/journals/bulletin_of_the_history_of_medicine/v079/79.2slater.html- Malaria Atlas Project
- Tree of Life Plasmodium
- PlasmoDBPlasmoDBPlasmoDB is a genomic database for Plasmodium....