
Action potential
Overview
Physiology
Physiology is the science of the function of living systems. This includes how organisms, organ systems, organs, cells, and bio-molecules carry out the chemical or physical functions that exist in a living system. The highest honor awarded in physiology is the Nobel Prize in Physiology or...
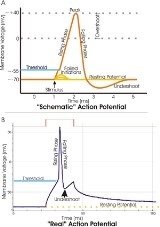
, an action potential is a short-lasting event in which the electrical membrane potential
Membrane potential
Membrane potential is the difference in electrical potential between the interior and exterior of a biological cell. All animal cells are surrounded by a plasma membrane composed of a lipid bilayer with a variety of types of proteins embedded in it...
of a cell
Cell (biology)
The cell is the basic structural and functional unit of all known living organisms. It is the smallest unit of life that is classified as a living thing, and is often called the building block of life. The Alberts text discusses how the "cellular building blocks" move to shape developing embryos....
rapidly rises and falls, following a consistent trajectory. Action potentials occur in several types of animal cells, called excitable cells
Membrane potential
Membrane potential is the difference in electrical potential between the interior and exterior of a biological cell. All animal cells are surrounded by a plasma membrane composed of a lipid bilayer with a variety of types of proteins embedded in it...
, which include neuron
Neuron
A neuron is an electrically excitable cell that processes and transmits information by electrical and chemical signaling. Chemical signaling occurs via synapses, specialized connections with other cells. Neurons connect to each other to form networks. Neurons are the core components of the nervous...
s, muscle cells
Myocyte
A myocyte is the type of cell found in muscles. They arise from myoblasts.Each myocyte contains myofibrils, which are long, long chains of sarcomeres, the contractile units of the cell....
, and endocrine cells, as well as in some plant cell
Plant cell
Plant cells are eukaryotic cells that differ in several key respects from the cells of other eukaryotic organisms. Their distinctive features include:...
s. In neurons, they play a central role in cell-to-cell communication. In other types of cells, their main function is to activate intracellular processes.
Unanswered Questions
Discussions