

Barrel cortex
Encyclopedia
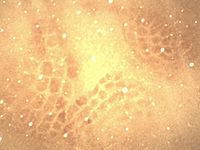
Neocortex
The neocortex , also called the neopallium and isocortex , is a part of the brain of mammals. It is the outer layer of the cerebral hemispheres, and made up of six layers, labelled I to VI...
where somatosensory inputs from the contralateral side of the body come in from the thalamus
Thalamus
The thalamus is a midline paired symmetrical structure within the brains of vertebrates, including humans. It is situated between the cerebral cortex and midbrain, both in terms of location and neurological connections...
. Barrels are found in some species of rodent
Rodent
Rodentia is an order of mammals also known as rodents, characterised by two continuously growing incisors in the upper and lower jaws which must be kept short by gnawing....
s and species of at least two other orders
Order (biology)
In scientific classification used in biology, the order is# a taxonomic rank used in the classification of organisms. Other well-known ranks are life, domain, kingdom, phylum, class, family, genus, and species, with order fitting in between class and family...
. The rest of this article will focus on the 'whisker barrels' in the somatosensory cortex of rodents. Inputs from the thalamus carrying information from a given whisker terminate in discrete areas of layer IV forming anatomically distinguishable areas (barrels) which are separated from each other by areas called septa. These structures were first discovered by Woolsey and Van der Loos (1970). Recognizing that the array was similar to that of the vibrissae on the mystacial pad, they hypothesized that the barrels were the "cortical correlates of the mystacial vibrissae" and that "one barrel represents one vibrissa".
Due to its distinctive cytoarchitectonics
Cytoarchitectonics of the cerebral cortex
The cytoarchitectonics of the cerebral cortex is the study of neuronal cell bodies cytoarchitecture in the cerebral cortex of the brain.-History:...
and functional significance, the barrel cortex has played an important role in neuroscience. The majority of what is known about corticothalamic processing comes from studying the barrel cortex and researchers have intensively studied the barrel cortex as a model of neocortical column.
Neurons within the layer IV barrel cortex directly code for whisker displacement. That is to say, that the neuron within a given barrel will fire when the whisker that barrel represents is moved. These neurons also show directional sensitivity, certain neurons will only fire when the whisker is moved in a specific direction.
Neurons in the barrel cortex exhibit the property of synaptic plasticity
Synaptic plasticity
In neuroscience, synaptic plasticity is the ability of the connection, or synapse, between two neurons to change in strength in response to either use or disuse of transmission over synaptic pathways. Plastic change also results from the alteration of the number of receptors located on a synapse...
that allows them to alter the vibrissae to which they respond depending on the rodent's history of tactile experience. The field of barrel cortex research has matured to the stage where a book has been published on the subject.
The Posteromedial Barrel Subfield

Afferent pathways
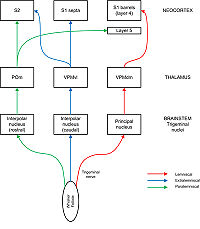
Trigeminal nerve
The trigeminal nerve contains both sensory and motor fibres. It is responsible for sensation in the face and certain motor functions such as biting, chewing, and swallowing. Sensory information from the face and body is processed by parallel pathways in the central nervous system...
carries afferent fibres from the follicles into the brainstem where they synapse with neurons in four different nuclei: principal, interpolar, oral, and caudal. The most important of these pathways for our purposes is the lemniscal pathway, in which axons from the principal nucleus cross over and project to “barreloids” in the dorsomedial section of the ventroposterior medial nucleus (VPMdm) of the thalamus. Neurons in VPMdm project mainly to barrels in layer 4 of primary somatosensory cortex (S1). In the extralemniscal pathway, neurons of the interpolar nucleus project to the ventrolateral section of the VPM (VPMvl). Neurons in VPMvl project to septa between the barrels and to secondary somatosensory cortex
Secondary somatosensory cortex
The human secondary somatosensory cortex is a region of cerebral cortex lying mostly on the parietal operculum.Region S2 was first described by Adrian in 1940, who found that feeling in cats' feet was not only represented in the previously described primary somatosensory cortex but also in a...
(S2). The paralemniscal pathway runs via the interpolar nucleus and the posterior nucleus (POm) of the thalamus to S2 and to diffuse targets in barrel cortex especially layer 5. These different pathways are thought to transmit different modalities of sensory information from the whisker.
Barrel cortex as an experimental system
The barrel cortex of the rodent has proved to be a fruitful model system for investigating plasticity in primary somatosensory cortex (S1). There are several reasons for its popularity (adapted from Feldman and Brecht, 2005 ):- The barrel cortex contains an orderly map which corresponds with the arrangement of the facial whiskers.
- The cortical representations of the whiskers correspond one-to-one with barrel structures which are visible under the microscope even in unstained slices.
- Barrel cortex is superficially located and so amenable to in vivo investigation.
- Sensory deprivation, by trimming or plucking of whiskers, or stimulation, by whisker vibration, is easy to arrange experimentally.
- The use of transgenic mice allows genetic and molecular mechanisms of plasticity to be investigated.
- Barrel columns are a specialized example of cortical columns, which are believed to be an important feature of cortical functional organization, suggesting that results from barrel cortex may be generally applicable to the rest of the sensory cortex.
Experience-dependent plasticity
Experience-dependent plasticity is commonly studied in the barrel cortex by partially depriving it of sensory input, either by lesioning elements of the afferent pathway (e.g. the trigeminal nerve) or by ablating, plucking, or trimming some of the facial whiskers. Only lesioning affects the anatomical structure of the barrels, but innocuous forms of deprivation can induce rapid changes in the cortical map into adulthood, without any corresponding changes in the barrel structures. These two paradigms seem to work by different mechanisms.Some forms of plasticity in the barrel cortex display a critical period. Plucking whiskers in neonatal rats causes a long-lasting expansion of the representation of the spared whisker in layer 4. However, layer 4 plasticity rapidly diminishes if sensory deprivation begins after day 4 of life (P4) whereas representations in layer 2/3 remain highly plastic into adulthood.
Two cortical processes run alongside each other when barrel cortex is deprived of sensory input from some whiskers to produce representational plasticity. In deprived cortex neuronal responses to spared whiskers are enhanced, and responses to deprived whiskers are weakened. These two processes have different time courses, with the weakening of deprived response preceding the strengthening of spared response, implying that they have different underlying mechanisms. These two effects combine to produce an expansion of the cortical representation of spared whiskers into the representation of adjacent deprived whiskers.
It is likely that several different mechanisms are involved in producing experience-dependent
plasticity in a whisker deprivation protocol (adapted from Feldman and Brecht, 2005 ):
- There is an immediate unmasking effect, where loss of input to a deprived barrel column leads to a loss of inhibitory firing in that column which unmasks horizontal excitatory connections from adjacent spared columns. This does not explain longer-lasting plastic changes as the unmasking would disappear immediately if the deprived input was reinstated (for example by allowing the whisker to regrow).
- The involvement of LTP- and LTD-like processes is suggested by the observation that plasticity can be compromised by interfering with expression of calmodulin-dependent protein kinase II (CaMKII) or cyclic-AMP response element binding protein (CREB) in transgenic mice. Spike timing rather than frequency may be an important factor. Associative LTP has been demonstrated at layer 4 to layer 2/3 synapses when the layer 4 neuron fires 0-15 ms before the layer 2/3 neuron, and LTD is observed when this timing order is reversed. Such mechanisms could act rapidly to produce plastic changes within hours or days.
- Sensory deprivation has been demonstrated to cause changes in synaptic dynamics such as EPSP amplitude and frequency. The net effect of these changes is to increase the proportion of synaptic input which layer 2/3 neurons in deprived barrels receive from spared barrels. These observations suggest that other, more specific, mechanisms besides LTP/LTD are at play in experience-dependent plasticity.
- It seems intuitively likely that structural changes at the level of axons, dendrite branches, and dendrite spines underlie some of the long-term plastic changes in the cortex. Axonal remodelling has been reported in lesion-induced plasticity but not (until now) in experience-dependent plasticity, but a recent study by Cheetham et al. found that whisker trimming produces targeted axonal remodelling in spared cortex. Dendritic branching is important during prenatal and neonatal development, is involved in plasticity induced by lesions, but is not involved in experience-dependent plasticity. In vivo two-photon microscopy reveals that dendritic spines in mouse barrel cortex are highly dynamic and subject to continuous turnover, and may be associated with formation or deletion of synapses. It is likely that spine turnover is necessary but not sufficient to produce experience-dependent plasticity, and other mechanisms such as axonal remodelling are also needed to explain features such as savings from prior experience.
External links
Research groups working on barrel cortex:- Barrel Group, Freiburg
- Barrel cortex group, Cardiff
- Finnerty lab, MRC Centre for Neurodegeneration Research, London
Books on barrel cortex
Barrel Cortex book