

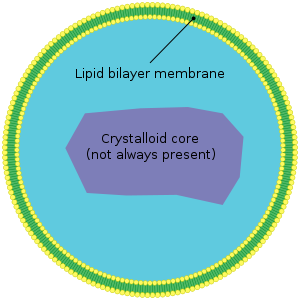
Peroxisome
Encyclopedia
Catabolism
Catabolism is the set of metabolic pathways that break down molecules into smaller units and release energy. In catabolism, large molecules such as polysaccharides, lipids, nucleic acids and proteins are broken down into smaller units such as monosaccharides, fatty acids, nucleotides, and amino...
of very long chain fatty acid
Very long chain fatty acid
A very long chain fatty acid is a fatty acid with aliphatic tails longer than 22 carbons.Unlike most fatty acids, VLCFAs are too long to be metabolized in the mitochondria, and must be metabolized in peroxisomes....
s, branched chain fatty acids, D-amino acids, polyamines, and biosynthesis of plasmalogens, etherphospholipids critical for the normal function of mammalian brains and lungs. They also contain approximately 10% of the total activity of two enzymes in the pentose phosphate pathway, which is important for energy metabolism. It is vigorously debated if peroxisomes are involved in isoprenoid and cholesterol
Cholesterol
Cholesterol is a complex isoprenoid. Specifically, it is a waxy steroid of fat that is produced in the liver or intestines. It is used to produce hormones and cell membranes and is transported in the blood plasma of all mammals. It is an essential structural component of mammalian cell membranes...
synthesis in animals. Other known peroxisomal functions include the glyoxylate cycle
Glyoxylate cycle
The glyoxylate cycle, a variation of the Tricarboxylic Acid Cycle, is an anabolic metabolic pathway occurring in plants, bacteria, protists, fungi and several microorganisms, such as E. coli and yeast. The glyoxylate cycle centers on the conversion of acetyl-CoA to succinate for the synthesis of...
in germinating seeds ("glyoxysomes"), photorespiration
Photorespiration
Photorespiration, or "'photo-respiration'", is a process in plant metabolism by which RuBP has oxygen added to it by the enzyme , instead of carbon dioxide during normal photosynthesis. This is the beginning step of the Calvin-Benson cycle...
in leaves, glycolysis
Glycolysis
Glycolysis is the metabolic pathway that converts glucose C6H12O6, into pyruvate, CH3COCOO− + H+...
in trypanosomes ("glycosome
Glycosome
The glycosome is a membrane-enclosed organelle that contains the glycolytic enzymes. It is found in a few species of protozoa, most notably in the human pathogenic trypanosomes, which can cause sleeping sickness and Chagas's disease, and Leishmania. The organelle is bounded by a single membrane and...
s"), and methanol
Methanol
Methanol, also known as methyl alcohol, wood alcohol, wood naphtha or wood spirits, is a chemical with the formula CH3OH . It is the simplest alcohol, and is a light, volatile, colorless, flammable liquid with a distinctive odor very similar to, but slightly sweeter than, ethanol...
and/or amine oxidation and assimilation in some yeasts.
Peroxisomes were identified as organelles by the Belgian cytologist Christian de Duve
Christian de Duve
Christian René, viscount de Duve is a Nobel Prize-winning cytologist and biochemist. De Duve was born in Thames Ditton, Surrey, Great Britain, as a son of Belgian refugees. They returned to Belgium in 1920...
in 1967 after they had been first described by a Swedish doctoral student, J. Rhodin in 1954.
Metabolic functions
A major function of the peroxisome is the breakdown of very long chain fatty acids through beta-oxidation. In animal cells, the very long fatty acids are converted to medium chain fatty acids, which are subsequently shuttled to mitochondria where they are eventually broken down to carbon dioxide and water. In yeast and plant cells, this process is exclusive for the peroxisome.The first reactions in the formation of plasmalogen
Plasmalogen
Plasmalogens are a type of ether phospholipid characterized by the presence of a vinyl ether linkage at the sn-1 position and an ester linkage at the sn-2 position . In mammals, the sn-1 position is typically derived from C16:0, C18:0, or C18:1 fatty alcohols while the sn-2 position is most...
in animal cells also occur in peroxisomes. Plasmalogen is the most abundant phospholipid in myelin
Myelin
Myelin is a dielectric material that forms a layer, the myelin sheath, usually around only the axon of a neuron. It is essential for the proper functioning of the nervous system. Myelin is an outgrowth of a type of glial cell. The production of the myelin sheath is called myelination...
. Deficiency of plasmalogens causes profound abnormalities in the myelination of nerve cells
Neuron
A neuron is an electrically excitable cell that processes and transmits information by electrical and chemical signaling. Chemical signaling occurs via synapses, specialized connections with other cells. Neurons connect to each other to form networks. Neurons are the core components of the nervous...
, which is one reason why many peroxisomal disorders affect the nervous system. Peroxisomes also play a role in the production of bile
Bile
Bile or gall is a bitter-tasting, dark green to yellowish brown fluid, produced by the liver of most vertebrates, that aids the process of digestion of lipids in the small intestine. In many species, bile is stored in the gallbladder and upon eating is discharged into the duodenum...
acids important for the absorption of fats and fat-soluble vitamins, such as vitamins A and K. Skin disorders are features of genetic disorders affecting peroxisome function as a result.
Peroxisomes contain oxidative enzyme
Enzyme
Enzymes are proteins that catalyze chemical reactions. In enzymatic reactions, the molecules at the beginning of the process, called substrates, are converted into different molecules, called products. Almost all chemical reactions in a biological cell need enzymes in order to occur at rates...
s, such as catalase
Catalase
Catalase is a common enzyme found in nearly all living organisms that are exposed to oxygen, where it catalyzes the decomposition of hydrogen peroxide to water and oxygen...
, D-amino acid oxidase
D-amino acid oxidase
D-amino acid oxidase is a peroxisomal enzyme containing FAD as cofactor that is expressed in a wide range of species from yeasts to human. It is not present in bacteria or in plants...
, and uric acid oxidase. However the last enzyme is absent in humans, explaining the disease known as gout
Gout
Gout is a medical condition usually characterized by recurrent attacks of acute inflammatory arthritis—a red, tender, hot, swollen joint. The metatarsal-phalangeal joint at the base of the big toe is the most commonly affected . However, it may also present as tophi, kidney stones, or urate...
, caused by the accumulation of uric acid. Certain enzymes within the peroxisome, by using molecular oxygen, remove hydrogen atoms from specific organic substrates (labeled as R), in an oxidative reaction, producing hydrogen peroxide
Hydrogen peroxide
Hydrogen peroxide is the simplest peroxide and an oxidizer. Hydrogen peroxide is a clear liquid, slightly more viscous than water. In dilute solution, it appears colorless. With its oxidizing properties, hydrogen peroxide is often used as a bleach or cleaning agent...
(H2O2, itself toxic):

Catalase
Catalase
Catalase is a common enzyme found in nearly all living organisms that are exposed to oxygen, where it catalyzes the decomposition of hydrogen peroxide to water and oxygen...
, another peroxisomal enzyme, uses this H2O2 to oxidize other substrates, including phenols
Phenols
In organic chemistry, phenols, sometimes called phenolics, are a class of chemical compounds consisting of a hydroxyl group bonded directly to an aromatic hydrocarbon group...
, formic acid
Formic acid
Formic acid is the simplest carboxylic acid. Its chemical formula is HCOOH or HCO2H. It is an important intermediate in chemical synthesis and occurs naturally, most notably in the venom of bee and ant stings. In fact, its name comes from the Latin word for ant, formica, referring to its early...
, formaldehyde
Formaldehyde
Formaldehyde is an organic compound with the formula CH2O. It is the simplest aldehyde, hence its systematic name methanal.Formaldehyde is a colorless gas with a characteristic pungent odor. It is an important precursor to many other chemical compounds, especially for polymers...
, and alcohol
Alcohol
In chemistry, an alcohol is an organic compound in which the hydroxy functional group is bound to a carbon atom. In particular, this carbon center should be saturated, having single bonds to three other atoms....
, by means of the peroxidation reaction:
 , thus eliminating the poisonous hydrogen peroxide in the process.
, thus eliminating the poisonous hydrogen peroxide in the process.This reaction is important in liver and kidney cells, where the peroxisomes detoxify various toxic substances that enter the blood. About 25% of the ethanol
Ethanol
Ethanol, also called ethyl alcohol, pure alcohol, grain alcohol, or drinking alcohol, is a volatile, flammable, colorless liquid. It is a psychoactive drug and one of the oldest recreational drugs. Best known as the type of alcohol found in alcoholic beverages, it is also used in thermometers, as a...
humans drink is oxidized to acetaldehyde
Acetaldehyde
Acetaldehyde is an organic chemical compound with the formula CH3CHO or MeCHO. It is one of the most important aldehydes, occurring widely in nature and being produced on a large scale industrially. Acetaldehyde occurs naturally in coffee, bread, and ripe fruit, and is produced by plants as part...
in this way. In addition, when excess H2O2 accumulates in the cell, catalase converts it to H2O through this reaction:
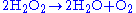
In higher plants,peroxisomes contain also a complex battery of antioxidative
enzymes such as superoxide dismutase, the components of the ascorbate-glutathione cycle, and the NADP-dehydrogenases of the pentose-phosphate pathway. It has
been demonstrated the generation of superoxide
Superoxide
A superoxide, also known by the obsolete name hyperoxide, is a compound that possesses the superoxide anion with the chemical formula O2−. The systematic name of the anion is dioxide. It is important as the product of the one-electron reduction of dioxygen O2, which occurs widely in nature...
(O2•-) and nitric oxide
Nitric oxide
Nitric oxide, also known as nitrogen monoxide, is a diatomic molecule with chemical formula NO. It is a free radical and is an important intermediate in the chemical industry...
(•NO) radicals.
The peroxisome of plant cells is polarised when fighting fungal penetration. Infection causes a glucosinolate
Glucosinolate
The glucosinolates are a class of organic compounds that contain sulfur and nitrogen and are derived from glucose and an amino acid. They occur as secondary metabolites of almost all plants of the order Brassicales , but also in the genus Drypetes .-Chemistry:Glucosinolates are water-soluble...
molecule to play an antifungal role to be made and delivered to the outside of the cell through the action of the peroxisomal proteins (PEN2 and PEN3).
Peroxisome assembly
Peroxisomes can be derived from the endoplasmic reticulumEndoplasmic reticulum
The endoplasmic reticulum is an organelle of cells in eukaryotic organisms that forms an interconnected network of tubules, vesicles, and cisternae...
and replicate by fission. Peroxisome matrix proteins are translated in the cytoplasm prior to import. Specific amino acid sequences (PTS or peroxisomal targeting signal
Peroxisomal targeting signal
A peroxisomal targeting signal is a region of the peroxisomal protein that receptors recognize and bind to.- Overview:All peroxisomal proteins are synthesized in the cytoplasm and must be directed to the peroxisome. The first step in this process is the binding of the protein to a receptor. The...
) at the C-terminus (PTS1) or N-terminus (PTS2) of peroxisomal matrix proteins signals them to be imported into the organelle. There are at least 32 known peroxisomal proteins, called peroxin
Peroxin
Peroxin is a protein found in peroxisomes.Deficiencies are associated with several peroxisomal disorders....
s, which participate in the process of peroxisome assembly. Proteins do not have to unfold to be imported into the peroxisome. The protein receptors, the peroxins PEX5
PEX5
Peroxisomal targeting signal 1 receptor is a protein that in humans is encoded by the PEX5 gene.-Interactions:PEX5 has been shown to interact with PEX12, PEX13 and PEX14.-External Links:* *...
and PEX7, accompany their cargoes (containing a PTS1 or a PTS5 amino acid sequence, respectively) all the way into the peroxisome where they release the cargo and then return to the cytosol
Cytosol
The cytosol or intracellular fluid is the liquid found inside cells, that is separated into compartments by membranes. For example, the mitochondrial matrix separates the mitochondrion into compartments....
- a step named recycling. A model describing the import cycle is referred to as the extended shuttle mechanism. There is now evidence that ATP hydrolysis is required for the recycling of receptors to the cytosol
Cytosol
The cytosol or intracellular fluid is the liquid found inside cells, that is separated into compartments by membranes. For example, the mitochondrial matrix separates the mitochondrion into compartments....
. Also, ubiquitination appears to be crucial for the export of PEX5 from the peroxisome, to the cytosol.
Associated medical conditions
Peroxisomal disorders are a class of medical conditions that typically affect the human nervous system as well as many other organ systems. Two common examples are X-linked adrenoleukodystrophyAdrenoleukodystrophy
Adrenoleukodystrophy is a rare, inherited disorder that leads to progressive brain damage, failure of the adrenal glands and eventually death. ALD is a disease in a group of genetic disorders called leukodystrophies, whose chief feature is damage to myelin...
and peroxisome biogenesis disorders.
Genes
PEX genes encode the protein machinery ("peroxins") required for proper peroxisome assembly, as described above. Membrane assembly and maintenance requires three of these (peroxins 3, 16, and 19) and may occur without the import of the matrix (lumen) enzymes. Proliferation of the organelle is regulated by Pex11p.Genes that encode peroxin proteins include: PEX1
PEX1
Peroxisome biogenesis factor 1, also known as PEX1, is a protein which in humans is encoded by the PEX1 gene.This gene encodes a member of the AAA protein family, a large group of ATPases associated with diverse cellular activities...
, PEX2 - PXMP3
PXMP3
Peroxisome assembly factor 1 is a protein that in humans is encoded by the PXMP3 gene.-External Links:* *...
, PEX3
PEX3
Peroxisomal biogenesis factor 3 is a protein that in humans is encoded by the PEX3 gene.-External Links:* *...
, PEX5
PEX5
Peroxisomal targeting signal 1 receptor is a protein that in humans is encoded by the PEX5 gene.-Interactions:PEX5 has been shown to interact with PEX12, PEX13 and PEX14.-External Links:* *...
, PEX6
PEX6
Peroxisome assembly factor 2 is a protein that in humans is encoded by the PEX6 gene.-Interactions:PEX6 has been shown to interact with PEX1 and PEX26.-External Links:* *...
, PEX7, PEX10
PEX10
Peroxisome biogenesis factor 10 is a protein that in humans is encoded by the PEX10 gene.- Function :Peroxisome biogenesis factor 10 os involved in import of peroxisomal matrix proteins. This protein localizes to the peroxisomal membrane...
, PEX11A
PEX11A
Peroxisomal membrane protein 11A is a protein that in humans is encoded by the PEX11A gene.-Further reading:...
, PEX11B
PEX11B
Peroxisomal membrane protein 11B is a protein that in humans is encoded by the PEX11B gene. It is involved in the regulation of peroxisome abundance.-Related gene problems:*TAR syndrome*1q21.1 deletion syndrome*1q21.1 duplication syndrome...
, PEX11G
PEX11G
PEX11G is a human gene that encodes the peroxisomal biogenesis factor 11 gamma for peroxisomes. It is located on chromosome 19....
, PEX12
PEX12
Peroxisome assembly protein 12 is a protein that in humans is encoded by the PEX12 gene.- Function :The peroxisome biogenesis disorders are a group of genetically heterogeneous diseases that are usually lethal in early infancy...
, PEX13
PEX13
Peroxisomal membrane protein PEX13 is a protein that in humans is encoded by the PEX13 gene.. It located on chromosome 2 next to KIAA1841-Interactions:PEX13 has been shown to interact with PEX14, PEX5 and PEX19.-External Links:* *...
, PEX14
PEX14
Peroxisomal membrane protein PEX14 is a protein that in humans is encoded by the PEX14 gene.-Interactions:PEX14 has been shown to interact with* PEX5* PEX7* PEX13-External Links:* *...
, PEX16
PEX16
Peroxisomal membrane protein PEX16 is a protein that in humans is encoded by the PEX16 gene.- Function :The protein encoded by this gene is an integral peroxisomal membrane protein. An inactivating nonsense mutation localized to this gene was observed in a patient with Zellweger syndrome of the...
, PEX19
PEX19
Peroxisomal biogenesis factor 19 is a protein that in humans is encoded by the PEX19 gene.-Interactions:PEX19 has been shown to interact with PEX10, PEX12, PEX13, ABCD3, ABCD1, PEX3, ABCD2, PEX11B and PEX16.-External Links:* *...
, PEX26
PEX26
Peroxisome assembly protein 26 is a protein that in humans is encoded by the PEX26 gene.-Interactions:PEX26 has been shown to interact with PEX1, PEX6 and SUFU.-External Links:* *...
, PEX28, PEX30, and PEX31
Evolutionary origins
The protein content of peroxisomes varies across species, but the presence of proteins common to many species has been used to suggest an endosymbioticEndosymbiotic theory
The endosymbiotic theory concerns the mitochondria, plastids , and possibly other organelles of eukaryotic cells. According to this theory, certain organelles originated as free-living bacteria that were taken inside another cell as endosymbionts...
origin; that is, peroxisomes evolved from bacteria that invaded larger cells as parasites, and very gradually evolved a symbiotic relationship. However, this view has been challenged by recent discoveries. For example, peroxisome-less mutants can restore peroxisomes upon introduction of the wild-type gene.
Two independent evolutionary analyses of the peroxisomal proteome
Proteome
The proteome is the entire set of proteins expressed by a genome, cell, tissue or organism. More specifically, it is the set of expressed proteins in a given type of cells or an organism at a given time under defined conditions. The term is a portmanteau of proteins and genome.The term has been...
found homologies between the peroxisomal import machinery and the ERAD pathway in the endoplasmic reticulum
Endoplasmic reticulum
The endoplasmic reticulum is an organelle of cells in eukaryotic organisms that forms an interconnected network of tubules, vesicles, and cisternae...
, along with a number of metabolic enzymes that were likely recruited from the mitochondria. Recently, it has been suggested that the peroxisome may have had an actinobacterial
Actinobacteria
Actinobacteria are a group of Gram-positive bacteria with high guanine and cytosine content. They can be terrestrial or aquatic. Actinobacteria is one of the dominant phyla of the bacteria....
origin, however, this is controversial.
Other related organelles
Other organelles of the microbodyMicrobody
A microbody is a cytoplasmic organelle of a more or less globular shape that comprises degradative enzymes bound within a single membrane. Microbodies are specialized as containers for metabolic activity.-Types:...
family related to peroxisomes include glyoxysome
Glyoxysome
Glyoxysomes are specialized peroxisomes found in plants and also in filamentous fungi....
s of plant
Plant
Plants are living organisms belonging to the kingdom Plantae. Precise definitions of the kingdom vary, but as the term is used here, plants include familiar organisms such as trees, flowers, herbs, bushes, grasses, vines, ferns, mosses, and green algae. The group is also called green plants or...
s and filamentous fungi, glycosome
Glycosome
The glycosome is a membrane-enclosed organelle that contains the glycolytic enzymes. It is found in a few species of protozoa, most notably in the human pathogenic trypanosomes, which can cause sleeping sickness and Chagas's disease, and Leishmania. The organelle is bounded by a single membrane and...
s of kinetoplastid
Kinetoplastid
The kinetoplastids are a group of single-cell flagellate protozoa, including a number of parasites responsible for serious diseases in humans and other animals, as well as various forms found in soil and aquatic environments...
s and Woronin bodies
Woronin body
A Woronin body is a peroxisome-derived, dense core microbody with a double membrane found near the septae that divide hyphal compartments in filamentous Ascomycota. One established function of Woronin bodies is the plugging of the septal pores after hyphal wounding, which restricts theloss of...
of filamentous fungi.