

Myosin
Encyclopedia
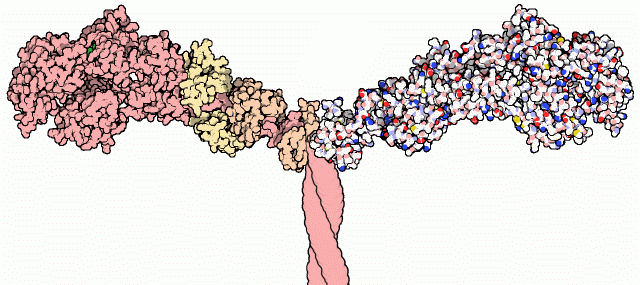
Muscle
Muscle is a contractile tissue of animals and is derived from the mesodermal layer of embryonic germ cells. Muscle cells contain contractile filaments that move past each other and change the size of the cell. They are classified as skeletal, cardiac, or smooth muscles. Their function is to...
contraction and their involvement in a wide range of other eukaryotic motility processes. They are responsible for actin
Actin
Actin is a globular, roughly 42-kDa moonlighting protein found in all eukaryotic cells where it may be present at concentrations of over 100 μM. It is also one of the most highly-conserved proteins, differing by no more than 20% in species as diverse as algae and humans...
-based motility
Motility
Motility is a biological term which refers to the ability to move spontaneously and actively, consuming energy in the process. Most animals are motile but the term applies to single-celled and simple multicellular organisms, as well as to some mechanisms of fluid flow in multicellular organs, in...
. The term was originally used to describe a group of similar ATPases found in striated and smooth muscle cells. Following the discovery by Pollard and Korn of enzymes with myosin-like function in Acanthamoeba castellanii
Acanthamoeba
Acanthamoeba is a genus of amoebae, one of the most common protozoa in soil, and also frequently found in fresh water and other habitats. The cells are small, usually 15 to 35 μm in length and oval to triangular in shape when moving. The pseudopods form a clear hemispherical lobe at the anterior,...
, a large number of divergent myosin genes have been discovered throughout eukaryotes. Thus, although myosin was originally thought to be restricted to muscle cells (hence, "myo"), there is no single "myosin" but rather a huge superfamily of genes whose protein products share the basic properties of actin binding, ATP hydrolysis (ATPase enzyme activity), and force transduction. Virtually all eukaryotic cells contain myosin isoforms. Some isoforms have specialized functions in certain cell types (such as muscle), while other isoforms are ubiquitous. The structure and function of myosin is strongly conserved across species, to the extent that rabbit muscle myosin II will bind to actin from an amoeba.
Domains
Most myosin molecules are composed of a headMyosin head
Muscle contraction is caused by the sliding action of the thick filaments over the thin filaments of the myofibril. Myosin is a major component of thick filaments and exists as a hexamer of 2 heavy chains, 2 alkali light chains, and 2 regulatory light chains...
, neck, and tail domain.
- The head domainMyosin headMuscle contraction is caused by the sliding action of the thick filaments over the thin filaments of the myofibril. Myosin is a major component of thick filaments and exists as a hexamer of 2 heavy chains, 2 alkali light chains, and 2 regulatory light chains...
binds the filamentous actinActinActin is a globular, roughly 42-kDa moonlighting protein found in all eukaryotic cells where it may be present at concentrations of over 100 μM. It is also one of the most highly-conserved proteins, differing by no more than 20% in species as diverse as algae and humans...
, and uses ATPAdenosine triphosphateAdenosine-5'-triphosphate is a multifunctional nucleoside triphosphate used in cells as a coenzyme. It is often called the "molecular unit of currency" of intracellular energy transfer. ATP transports chemical energy within cells for metabolism...
hydrolysisHydrolysisHydrolysis is a chemical reaction during which molecules of water are split into hydrogen cations and hydroxide anions in the process of a chemical mechanism. It is the type of reaction that is used to break down certain polymers, especially those made by condensation polymerization...
to generate force and to "walk" along the filament towards the barbed (+) end (with the exception of myosin VI, which moves towards the pointed (-) end). - the neck domain acts as a linker and as a lever arm for transducing force generated by the catalytic motor domain. The neck domain can also serve as a binding site for myosin light chains which are distinct proteins that form part of a macromolecular complex and generally have regulatory functions.
- The tail domain generally mediates interaction with cargo molecules and/or other myosin subunits. In some cases, the tail domain may play a role in regulating motor activity.
Power stroke
Multiple myosin II molecules generate force in skeletal muscleMuscle contraction
Muscle fiber generates tension through the action of actin and myosin cross-bridge cycling. While under tension, the muscle may lengthen, shorten, or remain the same...
through a power stroke mechanism fuelled by the energy released from ATP hydrolysis. The power stroke occurs at the release of phosphate from the myosin molecule after the ATP hydrolysis while myosin is tightly bound to actin. The effect of this release is a conformational change in the molecule that pulls against the actin. The release of the ADP molecule and binding of a new ATP molecule will release myosin from actin. ATP hydrolysis within the myosin will cause it to bind to actin again to repeat the cycle. The combined effect of the myriad power strokes causes the muscle to contract.
Nomenclature, evolution, and the family tree
The wide variety of myosin genes found throughout the eukaryotic phyla were named according to different schemes as they were discovered. The nomenclature can therefore be somewhat confusing when attempting to compare the functions of myosin proteins within and between organisms.Skeletal muscle myosin, the most conspicuous of the myosin superfamily due to its abundance in muscle fibers, was the first to be discovered. This protein makes up part of the sarcomere and forms macromolecular filaments composed of multiple myosin subunits. Similar filament-forming myosin proteins were found in cardiac muscle, smooth muscle, and non-muscle cells. However, beginning in the 1970s researchers began to discover new myosin genes in simple eukaryotes encoding proteins that acted as monomers and were therefore entitled Class I myosins. These new myosins were collectively termed "unconventional myosins" and have been found in many tissues other than muscle. These new superfamily members have been grouped according to phylogenetic relationships derived from a comparison of the amino acid sequences of their head domains, with each class being assigned a Roman numeral(see phylogenetic tree). The unconventional myosins also have divergent tail domains, suggesting unique functions. The now diverse array of myosins likely evolved from an ancestral precursor (see picture).

Human genome
The human genome is the genome of Homo sapiens, which is stored on 23 chromosome pairs plus the small mitochondrial DNA. 22 of the 23 chromosomes are autosomal chromosome pairs, while the remaining pair is sex-determining...
contains over 40 different myosin genes
Gênes
Gênes is the name of a département of the First French Empire in present Italy, named after the city of Genoa. It was formed in 1805, when Napoleon Bonaparte occupied the Republic of Genoa. Its capital was Genoa, and it was divided in the arrondissements of Genoa, Bobbio, Novi Ligure, Tortona and...
.
These differences in shape also determine the speed at which myosins can move along actin filaments. The hydrolysis of ATP and the subsequent release of the phosphate
Phosphate
A phosphate, an inorganic chemical, is a salt of phosphoric acid. In organic chemistry, a phosphate, or organophosphate, is an ester of phosphoric acid. Organic phosphates are important in biochemistry and biogeochemistry or ecology. Inorganic phosphates are mined to obtain phosphorus for use in...
group causes the "power stroke
Power stroke
Power Stroke may refer to:In motoring:*Power stroke , the stroke of a cyclic motor which generates force*Ford Power Stroke engine, Ford diesel engineOther:*In baseball, a batter who hits for extra bases is said to have a power stroke...
," in which the "lever arm" or "neck" region of the heavy chain is dragged forward. Since the power stroke always moves the lever arm by the same angle, the length of the lever arm determines the displacement of the cargo relative to the actin filament. A longer lever arm will cause the cargo to traverse a greater distance even though the lever arm undergoes the same angular displacement - just as a person with longer legs can move farther with each individual step. The velocity of a myosin motor depends upon the rate at which it passes through a complete kinetic cycle of ATP binding to the release of ADP.
Myosin I
Myosin I functions as monomer and functions in vesicleVesicle (biology)
A vesicle is a bubble of liquid within another liquid, a supramolecular assembly made up of many different molecules. More technically, a vesicle is a small membrane-enclosed sack that can store or transport substances. Vesicles can form naturally because of the properties of lipid membranes , or...
transport. It has step size of 10 nm and has been implicated as being responsible for the adaptation response of the stereocilia in the inner ear.
Myosin II
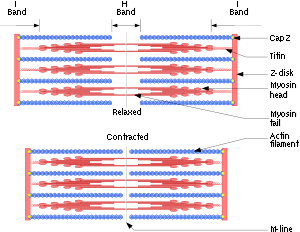
Muscle contraction
Muscle fiber generates tension through the action of actin and myosin cross-bridge cycling. While under tension, the muscle may lengthen, shorten, or remain the same...
in muscle cells.
- Myosin II contains two heavy chains, each about 2000 amino acids in length, which constitute the head and tail domains. Each of these heavy chains contains the N-terminal head domain, while the C-terminal tails take on a coiled-coil morphology, holding the two heavy chains together (imagine two snakes wrapped around each other, such as in a caduceusCaduceusThe caduceus is the staff carried by Hermes in Greek mythology. The same staff was also borne by heralds in general, for example by Iris, the messenger of Hera. It is a short staff entwined by two serpents, sometimes surmounted by wings...
). Thus, myosin II has two heads. The intermediate neck domain is the region creating the angle between the head and tail. In smooth muscle, there is a single gene (MYH11MYH11Myosin-11 is a protein that in humans is encoded by the MYH11 gene.- Function :Myosin-11 is a smooth muscle myosin belonging to the myosin heavy chain family...
) that codes for the heavy chains myosin II, but there are splice variantsAlternative splicingAlternative splicing is a process by which the exons of the RNA produced by transcription of a gene are reconnected in multiple ways during RNA splicing...
of this gene that result in four distinct isoforms. - It also contains 4 light chains, resulting in 2 per head, weighing 20 (MLC20) and 17 (MLC17) kDa. These bind the heavy chains in the "neck" region between the head and tail.
- The MLC20 is also known as the regulatory light chain and actively participates in muscle contractionMuscle contractionMuscle fiber generates tension through the action of actin and myosin cross-bridge cycling. While under tension, the muscle may lengthen, shorten, or remain the same...
. - The MLC17 is also known as the essential light chain. Its exact function is unclear, but it's believed that it contributes to the structural stability of the myosin head along with MLC20. Two variants of MLC17 (MLC17a/b) exist as a result of alternate splicing at the MLC17 gene.
- The MLC20 is also known as the regulatory light chain and actively participates in muscle contraction
In muscle cells, the long coiled-coil tails of the individual myosin molecules join together, forming the thick filaments of the sarcomere
Sarcomere
A sarcomere is the basic unit of a muscle. Muscles are composed of tubular muscle cells . Muscle cells are composed of tubular myofibrils. Myofibrils are composed of repeating sections of sarcomeres, which appear under the microscope as dark and light bands...
. The force-producing head domains stick out from the side of the thick filament, ready to walk along the adjacent actin-based thin filaments in response to the proper chemical signals.
Myosin III
Myosin III is a poorly understood member of the myosin family. It has been studied in vivoIn vivo
In vivo is experimentation using a whole, living organism as opposed to a partial or dead organism, or an in vitro controlled environment. Animal testing and clinical trials are two forms of in vivo research...
in the eyes of Drosophila
Drosophila
Drosophila is a genus of small flies, belonging to the family Drosophilidae, whose members are often called "fruit flies" or more appropriately pomace flies, vinegar flies, or wine flies, a reference to the characteristic of many species to linger around overripe or rotting fruit...
, where it is thought to play a role in phototransduction. A human homologue
Homology (biology)
Homology forms the basis of organization for comparative biology. In 1843, Richard Owen defined homology as "the same organ in different animals under every variety of form and function". Organs as different as a bat's wing, a seal's flipper, a cat's paw and a human hand have a common underlying...
gene for myosin III, MYO3A
MYO3A
Myosin IIIA is a protein that in humans is encoded by the MYO3A gene.-Further reading:...
, has been uncovered through the Human Genome Project
Human Genome Project
The Human Genome Project is an international scientific research project with a primary goal of determining the sequence of chemical base pairs which make up DNA, and of identifying and mapping the approximately 20,000–25,000 genes of the human genome from both a physical and functional...
and is expressed in the retina
Retina
The vertebrate retina is a light-sensitive tissue lining the inner surface of the eye. The optics of the eye create an image of the visual world on the retina, which serves much the same function as the film in a camera. Light striking the retina initiates a cascade of chemical and electrical...
and cochlea
Cochlea
The cochlea is the auditory portion of the inner ear. It is a spiral-shaped cavity in the bony labyrinth, making 2.5 turns around its axis, the modiolus....
.
Myosin IV
Myosin IV has a single IQ motif and a tail that lacks any coiled-coil forming sequence. It has homology similar to the tail domains of Myosin VII and XV.Myosin V
Myosin V is an unconventional myosin motor, which is functional as a dimer. It translocates (walks) along actin filaments traveling towards the barbed end (+ end) of the filaments. Myosin V was thought to be critical in vesicle movement from the center of the cell to the periphery, but has been shown to be more like a dynamic tether retaining vesicles and organelles in the actin-rich periphery of cells.Myosin VI
Myosin VI is an unconventional myosin motor, which is primarily functional as a dimer but also acts as a nonprocessive monomer. It walks along actin filaments travelling towards the minus end of the filaments. Myosin VI is thought to transport endocyticEndocytic cycle
Most animal cells take up portions of their surface plasma membranes in a process called endocytosis. The main route of endocytosis is the coated pit, which buds into a cell to form a cytoplasmic vesicle — a clathrin-coated vesicle. The membrane so internalised is processed in a series of...
vesicles into the cell.
Myosin VII
Myosin VII is an unconventional myosin with two FERM domainsFERM domain
In molecular biology, the FERM domain is a widespread protein module involved in localising proteins to the plasma membrane. FERM domains are found in a number of cytoskeletal-associated proteins that associate with various proteins at the interface between the plasma membrane and the cytoskeleton...
in the tail region. It has a very short coiled-coiled region. Myosin VII is required for phagocytosis in Dictyostelium discoideum
Dictyostelium discoideum
Dictyostelium discoideum is a species of soil-living amoeba belonging to the phylum Mycetozoa. D. discoideum, commonly referred to as slime mold, is a eukaryote that transitions from a collection of unicellular amoebae into a multicellular slug and then into a fruiting body within its lifetime. D...
, spermatogenesis
Spermatogenesis
Spermatogenesis is the process by which male primary germ cells undergo division, and produce a number of cells termed spermatogonia, from which the primary spermatocytes are derived. Each primary spermatocyte divides into two secondary spermatocytes, and each secondary spermatocyte into two...
in C. elegans
Caenorhabditis elegans
Caenorhabditis elegans is a free-living, transparent nematode , about 1 mm in length, which lives in temperate soil environments. Research into the molecular and developmental biology of C. elegans was begun in 1974 by Sydney Brenner and it has since been used extensively as a model...
and stereocilia
Stereocilia
In the inner ear, stereocilia are the mechanosensing organelles of hair cells, which respond to fluid motion in numerous types of animals for various functions, including hearing and balance. They are about 10–50 micrometers in length and share some similar features of microvilli...
formation in mice and zebrafish.
Myosin VIII
Myosin VIII is a plant-specific myosin linked to cell division; specifically, it is involved in regulating the flow of cytoplasm between cells and in the localisation of vesicles to the phragmoplastPhragmoplast
thumb|300px|Phragmoplast and cell plate formation in a plant cell during cytokinesis. Left side: Phragmoplast forms and cell plate starts to assemble in the center of the cell. Towards the right: Phragmoplast enlarges in a donut-shape towards the outside of the cell, leaving behind mature cell...
.
Myosin IX
Myosin IX is a single-headed, minus-end directed group of motor proteins. The movement mechanism for this myosin is poorly understood.Myosin X
Myosin X is an unconventional myosin motor, which is functional as a dimer. In mammalian cells the motor is found to localize to filopodiaFilopodia
Filopodia are slender cytoplasmic projections that extend beyond the leading edge of lamellipodia in migrating cells. They contain actin filaments cross-linked into bundles by actin-binding proteins, e.g. fascin and fimbrin. Filopodia form focal adhesions with the substratum, linking it to the...
. Myosin X walks towards the barbed ends of filaments. Some research suggests that it preferentially walks on bundles of actin, rather than single filaments. It is the first myosin motor to exhibit this behavior.
Myosin XI
Myosin XI directs the movement of organelles as plastids and mitochondria in plant cells. It is responsible of the light-directed movement of chloroplasts according to light intensity and the formation of stromuleStromule
Stromules are microscopic structures found in plant cells. Stromules are highly dynamic structures extending from the surface of all plastid types, including proplastids, chloroplasts, etioplasts, leucoplasts, amyloplasts, and chromoplasts...
s interconnecting different plastids.
Myosin XIV
This myosin group has been found in the ApicomplexaApicomplexa
The Apicomplexa are a large group of protists, most of which possess a unique organelle called apicoplast and an apical complex structure involved in penetrating a host's cell. They are unicellular, spore-forming, and exclusively parasites of animals. Motile structures such as flagella or...
phylum. The myosins localise to plasma membranes of the intracellular parasites
Parasitism
Parasitism is a type of symbiotic relationship between organisms of different species where one organism, the parasite, benefits at the expense of the other, the host. Traditionally parasite referred to organisms with lifestages that needed more than one host . These are now called macroparasites...
and may then be involved in the cell invasion process.
This myosin is also found in the ciliated protozoan Tetrahymena thermaphila. Known functions include: transporting phagosomes to the nucleus and perturbing the developmentally regulated elimination of the macronucleus during conjugation.
Myosin XV
Myosin XV is a motor that is involved in the structuring and motion of stereociliaStereocilia
In the inner ear, stereocilia are the mechanosensing organelles of hair cells, which respond to fluid motion in numerous types of animals for various functions, including hearing and balance. They are about 10–50 micrometers in length and share some similar features of microvilli...
of in the inner ear. It is thought to be functional as a monomer.
Genes in humans
Note that not all of these genes are active.- Class I: MYO1AMYO1AMyosin-Ia is a protein that in humans is encoded by the MYO1A gene.The protein encoded by this gene belongs to the myosin superfamily. Myosins are molecular motors that, upon interaction with actin filaments, utilize energy from ATP hydrolysis to generate mechanical force. Each myosin has a...
, MYO1BMYO1BMyosin-Ib is a protein that in humans is encoded by the MYO1B gene.-Further reading:...
, MYO1CMYO1CMyosin-Ic is a protein that in humans is encoded by the MYO1C gene.-Further reading:...
, MYO1D, MYO1EMYO1EMyosin-Ie is a protein that in humans is encoded by the MYO1E gene.-Further reading:...
, MYO1FMYO1FMyosin-If is a protein that in humans is encoded by the MYO1F gene.-Further reading:...
, MYO1GMyo1GMyosin 1G or Myo1G is a member of class I unconventional myosins. Its expression is highly restricted to hematopoietic tissues and cells. It localises exclusively to the plasma membrane and is dependent on both the motor domain and the tail domain. Myo1G regulates cell elasticity possibly by...
, MYO1H - Class II: MYH1MYH1Myosin-1 is a protein that in humans is encoded by the MYH1 gene.-Further reading:...
, MYH2MYH2Myosin-2 is a protein that in humans is encoded by the MYH2 gene.-Further reading:...
, MYH3MYH3Myosin-3 is a protein that in humans is encoded by the MYH3 gene.-Further reading:...
, MYH4MYH4Myosin-4 also known as myosin, heavy chain 4 is a protein which in humans is encoded by the MYH4 gene.-Further reading:...
, MYH6MYH6Myosin-6 is a protein that in humans is encoded by the MYH6 gene.- Function :Cardiac muscle myosin is a hexamer consisting of two heavy chain subunits, two light chain subunits, and two regulatory subunits. This gene encodes the alpha heavy chain subunit of cardiac myosin...
, MYH7MYH7MYH7 is a gene encoding a myosin heavy chain beta isoform expressed primarily in the heart.Changes in the relative abundance of MHC-β and MHC-α correlate with the contractile velocity of cardiac muscle...
, MYH7BMYH7BMyosin-7B also known as myosin, heavy chain 7B is a protein that in humans is encoded by the MYH7B gene....
, MYH8MYH8Myosin-8 is a protein that in humans is encoded by the MYH8 gene.-Further reading:...
, MYH9MYH9Myosin, heavy chain 9, non-muscle is a protein which in humans is encoded by the MYH9 gene.-Clinical significance:MYH9 polymorphisms have been shown to associate with glomerulosclerosis and non-diabetic end stage renal disease in African Americans and in Hispanic Americans, though it was later...
, MYH10MYH10Myosin-10 is a protein that in humans is encoded by the MYH10 gene.-Further reading:...
, MYH11MYH11Myosin-11 is a protein that in humans is encoded by the MYH11 gene.- Function :Myosin-11 is a smooth muscle myosin belonging to the myosin heavy chain family...
, MYH13MYH13Myosin-13 also known as myosin, heavy chain 13 is a protein which in humans is encoded by the MYH13 gene.- Function :MYH13 is a myosin whose expression is restricted primarily to the extrinsic eye muscles which are specialized for function in eye movement....
, MYH14MYH14Myosin-14 is a protein that in humans is encoded by the MYH14 gene.-Further reading:...
, MYH15MYH15Myosin-15 also known as myosin, heavy chain 15 is a protein that in humans is encoded by the MYH15 gene....
, MYH16 - Class III: MYO3AMYO3AMyosin IIIA is a protein that in humans is encoded by the MYO3A gene.-Further reading:...
, MYO3B - Class V: MYO5AMYO5AMyosin-Va is a protein that in humans is encoded by the MYO5A gene.-Interactions:MYO5A has been shown to interact with DYNLL1, RAB27A and DYNLL2.-Clinical significance:...
, MYO5BMYO5BMyosin-Vb is a protein that in humans is encoded by the MYO5B gene.Recent evidence suggests that Myosin VB is related to the creation of memories.Mutations of MYO5B causes microvillous inclusion disease.-Interactions:...
, MYO5C - Class VI: MYO6MYO6Myosin VI, also known as MYO6, is a protein. It has been found in humans, mice, fruit flies , and nematodes .-Interactions:MYO6 has been shown to interact with GIPC1 and DAB2....
- Class VII: MYO7AMYO7AMyosin VIIA is protein that in humans is encoded by the MYO7A gene. Myosin VIIA is a member of the unconventional myosin superfamily of proteins...
, MYO7B - Class IX: MYO9A, MYO9BMYO9BMyosin-IXb is a protein that in humans is encoded by the MYO9B gene.-Further reading:...
- Class X: MYO10MYO10Myosin X, also known as MYO10, is a protein which in humans is encoded by the MYO10 gene.Myosin X is an actin-based motor protein known to associate at the tips of filopodia.-Further reading:...
- Class XV: MYO15AMYO15AMyosin-XV is a protein that in humans is encoded by the MYO15A gene.-Further reading:...
- Class XVIII: MYO18AMYO18AMyosin-XVIIIa is a protein that in humans is encoded by the MYO18A gene.-Further reading:...
, MYO18BMYO18BMyosin-XVIIIb is a protein that in humans is encoded by the MYO18B gene.-Further reading:...
Myosin light chains are distinct and have their own properties. They are not considered "myosins" but are components of the macromolecular complexes that make up the functional myosin enzymes.
- Light chain: MYL1MYL1Myosin light chain 3, skeletal muscle isoform is a protein that in humans is encoded by the MYL1 gene.-Further reading:...
, MYL2MYL2Myosin regulatory light chain 2, ventricular/cardiac muscle isoform is a protein that in humans is encoded by the MYL2 gene.- External links :* * Info with links in the...
, MYL3MYL3Myosin light chain 3 is a protein that in humans is encoded by the MYL3 gene.- External links :*...
, MYL4MYL4Myosin light chain 4 is a protein that in humans is encoded by the MYL4 gene.-Further reading:...
, MYL5, MYL6MYL6Myosin light polypeptide 6 is a protein that in humans is encoded by the MYL6 gene.-Further reading:...
, MYL6BMYL6BMyosin light chain 6B is a protein that in humans is encoded by the MYL6B gene.-Further reading:...
, MYL7MYL7Myosin regulatory light chain 2, atrial isoform is a protein that in humans is encoded by the MYL7 gene.-Further reading:...
, MYL9MYL9Myosin regulatory light polypeptide 9 is a protein that in humans is encoded by the MYL9 gene.- Function :Myosin, a structural component of muscle, consists of two heavy chains and four light chains. The protein encoded by this gene is a myosin light chain that may regulate muscle contraction by...
, MYLIPMYLIPMyosin regulatory light chain interacting protein, also known as MYLIP, is a protein that in humans is encoded by the MYLIP gene.MYLIP is also known as IDOL "Inducible Degrader of the LDL receptor" based on its involvement in cholesterol regulation...
, MYLKMYLKMyosin light chain kinase, smooth muscle also known as kinase-related protein or telokin is an enzyme that in humans is encoded by the MYLK gene.- Function :...
, MYLK2MYLK2Myosin light chain kinase 2 also known as MYLK2 is an enzyme which in humans is encoded by the MYLK2 gene.- Function :This gene encodes a myosin light chain kinase, a calcium / calmodulin dependent enzyme, that is exclusively expressed in adult skeletal muscle....
, MYLL1
Paramyosin
Paramyosin is a large 93-115kDa muscleMuscle
Muscle is a contractile tissue of animals and is derived from the mesodermal layer of embryonic germ cells. Muscle cells contain contractile filaments that move past each other and change the size of the cell. They are classified as skeletal, cardiac, or smooth muscles. Their function is to...
protein
Protein
Proteins are biochemical compounds consisting of one or more polypeptides typically folded into a globular or fibrous form, facilitating a biological function. A polypeptide is a single linear polymer chain of amino acids bonded together by peptide bonds between the carboxyl and amino groups of...
that has been described in a number of diverse invertebrate
Invertebrate
An invertebrate is an animal without a backbone. The group includes 97% of all animal species – all animals except those in the chordate subphylum Vertebrata .Invertebrates form a paraphyletic group...
phyla. It is thought that invertebrate thick filaments are composted of an inner paramyosin core that is surrounded by myosin. The myosin interacts with actin
Actin
Actin is a globular, roughly 42-kDa moonlighting protein found in all eukaryotic cells where it may be present at concentrations of over 100 μM. It is also one of the most highly-conserved proteins, differing by no more than 20% in species as diverse as algae and humans...
resulting in fibre contraction. Paramyosin is found in many different invertebrate species, for example, Brachiopoda, Sipunculidea
Sipunculidea
Sipunculidea is a class of marine peanut worms. The tentacles form a circle around the mouth whereas those of the sister taxon, Phascolosomatidea, are only found above the mouth. Most species burrow in the substrate but some live in the empty shells of gastropods....
, Nematoda, Annelida, Mollusca
Mollusca
The Mollusca , common name molluscs or mollusksSpelled mollusks in the USA, see reasons given in Rosenberg's ; for the spelling mollusc see the reasons given by , is a large phylum of invertebrate animals. There are around 85,000 recognized extant species of molluscs. Mollusca is the largest...
, Arachnida, Insecta. Paramyosin is responsible for the "catch" mechanism that enables sustained contraction of muscles with very little energy expenditure, such that a clam
Clam
The word "clam" can be applied to freshwater mussels, and other freshwater bivalves, as well as marine bivalves.In the United States, "clam" can be used in several different ways: one, as a general term covering all bivalve molluscs...
can remain closed for extended periods.
Further reading
- Molecular Biology of the Cell. Alberts, Johnson, Lewis, Raff, Roberts, and Walter. 4th Edition. 949-952.
External links
- Myosin Video A video of a moving myosin motor protein.
- The Myosin Homepage
http://cellimages.ascb.org/cdm4/item_viewer.php?CISOROOT=/p4041coll12&CISOPTR=101&CISOBOX=1&REC=2 Animation of a moving myosin motor protein