

Microfilament
Encyclopedia
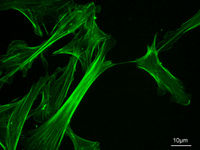
Cytoskeleton
The cytoskeleton is a cellular "scaffolding" or "skeleton" contained within a cell's cytoplasm and is made out of protein. The cytoskeleton is present in all cells; it was once thought to be unique to eukaryotes, but recent research has identified the prokaryotic cytoskeleton...
, a structure found in the cytoplasm
Cytoplasm
The cytoplasm is a small gel-like substance residing between the cell membrane holding all the cell's internal sub-structures , except for the nucleus. All the contents of the cells of prokaryote organisms are contained within the cytoplasm...
of all eukaryotic cells
Cell (biology)
The cell is the basic structural and functional unit of all known living organisms. It is the smallest unit of life that is classified as a living thing, and is often called the building block of life. The Alberts text discusses how the "cellular building blocks" move to shape developing embryos....
. These linear polymer
Biopolymer
Biopolymers are polymers produced by living organisms. Since they are polymers, Biopolymers contain monomeric units that are covalently bonded to form larger structures. There are three main classes of biopolymers based on the differing monomeric units used and the structure of the biopolymer formed...
s of actin
Actin
Actin is a globular, roughly 42-kDa moonlighting protein found in all eukaryotic cells where it may be present at concentrations of over 100 μM. It is also one of the most highly-conserved proteins, differing by no more than 20% in species as diverse as algae and humans...
subunits are flexible and relatively strong, resisting buckling by multi-piconewton compressive forces and filament fracture by nanonewton tensile forces. Microfilaments are highly versatile, functioning in cell crawling, amoeboid
Amoeboid
Amoeboids are single-celled life-forms characterized by an irregular shape."Amoeboid" and "amœba" are often used interchangeably even by biologists, and especially refer to a creature moving by using pseudopodia. Most references to "amoebas" or "amoebae" are to amoeboids in general rather than to...
movement, and changes in cell shape. In inducing this cell motility, one end of the actin filament elongates while the other end contracts, presumably by myosin II molecular motors. Additionally, they function as part of actomyosin-driven contractile molecular motors, wherein the thin filaments serve as tensile platforms for myosin's ATP-dependent pulling action in muscle contraction
Muscle contraction
Muscle fiber generates tension through the action of actin and myosin cross-bridge cycling. While under tension, the muscle may lengthen, shorten, or remain the same...
and uropod advancement.
Organization
Actin filaments are assembled in two general types of structures: bundles and networks. Bundles can be composed of polar filament arrays, in which all barbed ends point to the same end of the bundle, or non-polar arrays, where the barbed ends point towards both ends. A class of actin-binding proteinActin-binding protein
Actin-binding proteins are proteins that bind to actin. This may mean ability to bind actin monomers, or polymers, or both....
s, called cross-linking proteins, dictate the formation of these structures. Cross-linking proteins determine filament orientation and spacing in the bundles and networks. These structures are regulated by many other classes of actin-binding proteins, including motor proteins, branching proteins, severing proteins, polymerization promoters, and capping proteins.
In vitro self-assembly
Measuring approximately 6 nm in diameterDiameter
In geometry, a diameter of a circle is any straight line segment that passes through the center of the circle and whose endpoints are on the circle. The diameters are the longest chords of the circle...
, microfilaments are the thinnest fibers of the cytoskeleton. They are polymers of actin
Actin
Actin is a globular, roughly 42-kDa moonlighting protein found in all eukaryotic cells where it may be present at concentrations of over 100 μM. It is also one of the most highly-conserved proteins, differing by no more than 20% in species as diverse as algae and humans...
subunits (globular actin, or G-actin), which as part of the fiber are referred to as filamentous actin, or F-actin. Each microfilament is made up of two helix, interlaced strands of subunits. Much like microtubule
Microtubule
Microtubules are a component of the cytoskeleton. These rope-like polymers of tubulin can grow as long as 25 micrometers and are highly dynamic. The outer diameter of microtubule is about 25 nm. Microtubules are important for maintaining cell structure, providing platforms for intracellular...
s, actin filaments are polarized. Electron micrographs have provided evidence of their fast-growing barbed-ends and their slow-growing pointed-end. This polarity has been determined by the pattern created by the binding
Binding (molecular)
Molecular binding is an attractive interaction between two molecules which results in a stable association in which the molecules are in close proximity to each other...
of myosin S1 fragments: they themselves subunits of the larger myosin II protein complex. The pointed end is commonly referred to as the minus (-) end and the barbed end is referred to as the plus (+) end.
In vitro actin polymerization, or nucleation
Nucleation
Nucleation is the extremely localized budding of a distinct thermodynamic phase. Some examples of phases that may form by way of nucleation in liquids are gaseous bubbles, crystals or glassy regions. Creation of liquid droplets in saturated vapor is also characterized by nucleation...
, starts with the self-association of three G-actin monomers to form a trimer
Trimer (biochemistry)
In biochemistry, a trimer is a macromolecular complex formed by three, usually non-covalently bound, macromolecules like proteins or nucleic acids. A homo-trimer would be formed by three identical molecules. A hetero-trimer would be formed by three different macromolecules. Collagen is an example...
. ATP
Adenosine triphosphate
Adenosine-5'-triphosphate is a multifunctional nucleoside triphosphate used in cells as a coenzyme. It is often called the "molecular unit of currency" of intracellular energy transfer. ATP transports chemical energy within cells for metabolism...
-bound actin then itself binds the barbed end, and the ATP is subsequently hydrolyzed
Hydrolysis
Hydrolysis is a chemical reaction during which molecules of water are split into hydrogen cations and hydroxide anions in the process of a chemical mechanism. It is the type of reaction that is used to break down certain polymers, especially those made by condensation polymerization...
. ATP hydrolysis occurs with a half time
Half time (physics)
The half time is the time taken by a quantity to reach one half of its final value, where the rate of change is proportional to the difference between the present value and the final value....
of about 2 seconds, while the half time for the dissociation of the inorganic phosphate is about 6 minutes,. This autocatalyzed
Autocatalysis
A single chemical reaction is said to have undergone autocatalysis, or be autocatalytic, if the reaction product itself is the catalyst for that reaction....
event reduces the binding strength between neighboring subunits, and thus generally destabilizes the filament. In vivo
In vivo
In vivo is experimentation using a whole, living organism as opposed to a partial or dead organism, or an in vitro controlled environment. Animal testing and clinical trials are two forms of in vivo research...
actin polymerization is catalyzed by a new class of filament end-tracking molecular motors known as actoclampins. Recent evidence suggests that the rate of ATP hydrolysis and the rate of monomer incorporation are strongly coupled.
Subsequently, ADP
Adenosine diphosphate
Adenosine diphosphate, abbreviated ADP, is a nucleoside diphosphate. It is an ester of pyrophosphoric acid with the nucleoside adenosine. ADP consists of the pyrophosphate group, the pentose sugar ribose, and the nucleobase adenine....
-actin dissociates slowly from the pointed end, a process significantly accelerated by the actin-binding protein, cofilin
Cofilin
ADF/cofilin is a family of actin-binding proteins which disassembles actin filaments.Three highly conserved and highly identical genes belonging to this family have been described in human and mice:...
. ADP bound cofilin severs ADP-rich regions nearest the (–)-ends. Upon release, the free actin monomer slowly dissociates from ADP, which in turn rapidly binds to the free ATP diffusing
Diffusion
Molecular diffusion, often called simply diffusion, is the thermal motion of all particles at temperatures above absolute zero. The rate of this movement is a function of temperature, viscosity of the fluid and the size of the particles...
in the cytosol
Cytosol
The cytosol or intracellular fluid is the liquid found inside cells, that is separated into compartments by membranes. For example, the mitochondrial matrix separates the mitochondrion into compartments....
, thereby forming the ATP-actin monomeric units needed for further barbed-end filament elongation. This rapid turnover is important for the cell's movement. End-capping proteins such as CapZ
CapZ
CapZ is a protein that caps the barbed end of actin filaments in muscle cells. It is located in the muscle Z band....
prevent the addition or loss of monomers at the filament end where actin turnover is unfavorable, such as in the muscle apparatus.
Mechanism of force generation
As a result of ATP hydrolysis, filaments elongate approximately 10 times faster at their barbed ends than their pointed ends. At steady-state, the polymerization rate at the barbed end matches the depolymerization rate at the pointed end, and microfilaments are said to be treadmillingTreadmilling
Treadmilling is a phenomenon observed in many cellular cytoskeletal filaments, especially in actin filaments and microtubules. It occurs when one end of a filament grows in length while the other end shrinks resulting in a section of filament seemingly "moving" across a stratum or the cytosol...
. Treadmilling results in elongation in the barbed end and shortening in the pointed-end, so that the filament in total moves. Since both processes are energetically favorable, this means force is generated, the energy ultimately coming from the hydrolysis of this molecular unit of energy currrency
Adenosine triphosphate
Adenosine-5'-triphosphate is a multifunctional nucleoside triphosphate used in cells as a coenzyme. It is often called the "molecular unit of currency" of intracellular energy transfer. ATP transports chemical energy within cells for metabolism...
.
Actin in cells
Intracellular acting cytoskeletal assembly and disassembly are tightly regulated by cell signaling mechanisms. Many signal transductionSignal transduction
Signal transduction occurs when an extracellular signaling molecule activates a cell surface receptor. In turn, this receptor alters intracellular molecules creating a response...
systems use the actin cytoskeleton as a scaffold, holding them at or near the inner face of the peripheral membrane
Lipid bilayer
The lipid bilayer is a thin membrane made of two layers of lipid molecules. These membranes are flat sheets that form a continuous barrier around cells. The cell membrane of almost all living organisms and many viruses are made of a lipid bilayer, as are the membranes surrounding the cell nucleus...
. This subcellular location allows immediate and exquisite responsiveness to transmembrane receptor action and the resulting cascade of signal-processing enzymes. Because actin monomers must be recycled to sustain high rates of actin-based motility during chemotaxis
Chemotaxis
Chemotaxis is the phenomenon in which somatic cells, bacteria, and other single-cell or multicellular organisms direct their movements according to certain chemicals in their environment. This is important for bacteria to find food by swimming towards the highest concentration of food molecules,...
, cell signalling is believed to activate cofilin, the actin-filament depolymerizing protein which, as stated, binds to ADP-rich actin subunits nearest the filament's pointed-end and promotes filament fragmentation, with concomitant depolymerization in order to liberate actin monomers. The protein profilin
Profilin
Profilin is an actin-binding protein involved in the dynamic turnover and restructuring of the actin cytoskeleton. It is found in all eukaryotic organisms in most cells. Profilin is important for spatially and temporally controlled growth of actin microfilaments, which is an essential process in...
enhances the ability of monomers to assemble by stimulating the exchange of actin-bound ADP for solution-phase ATP to yield Actin-ATP and ADP. In most animal cells, monomeric actin is bound to profilin and thymosin-beta4, both of which preferentially bind with one-to-one stoichiometry to ATP-containing monomers. Although thymosin-beta4 is strictly a monomer-sequestering protein, the behavior of profilin is far more complex. Profilin is transferred to the leading edge by virtue of its PIP2 binding site, and it employs its poly-L-proline binding site to dock onto end-tracking proteins. Once bound, Profilin-Actin-ATP is loaded into the monomer-insertion site of actoclampin motors. Another important component in filament formation is the Arp2/3 complex
Arp2/3 complex
Arp2/3 complex is a seven-subunit protein that plays a major role in the regulation of the actin cytoskeleton. It is a major component of the actin cytoskeleton and is found in most in actin cytoskeleton-containing eukaryotic cells....
, which binds to the side of an already existing filament (or "mother filament"), where it nucleates the formation of a new daughter filament at a 70 degree angle relative to the mother filament, effecting a fan-like branched filament network.
Microfilament associated proteins
In non-muscle cells, actin filaments are formed proximal to membrane surfaces. Their formation and turnover are regulated by many proteins, including:- Filament end-tracking protein (e.g., forminsForminsFormins are a group of proteins that are involved in the polymerization of actin and associate with the fast-growing end of actin filaments. Most formins are Rho-GTPase effector proteins...
, VASPVASPViação Aérea São Paulo S/A, or VASP, was an airline with its head office in the VASP Building on the grounds of Congonhas-São Paulo Airport in São Paulo, Brazil...
, N-WASP) - Filament-nucleator known as the Actin-Related Protein-2/3 (or Arp2/3) complex
- Filament cross-linkers (e.g., α-actinin and fascinFascinFascin is a actin cross-linking protein.It is a 54-58 kilodalton monomeric actin filament bundling protein originally isolated from sea urchin egg but also found in Drosophila and vertebrates, including humans. Fascin is spaced at 11 nanometre intervals along the filament...
) - Actin monomer-binding proteins profilinProfilinProfilin is an actin-binding protein involved in the dynamic turnover and restructuring of the actin cytoskeleton. It is found in all eukaryotic organisms in most cells. Profilin is important for spatially and temporally controlled growth of actin microfilaments, which is an essential process in...
and thymosin-β4 - Filament barbed-end cappers such as Capping Protein and CapG, etc.
- Filament-severing proteins like gelsolinGelsolinGelsolin is an actin-binding protein that is a key regulator of actin filament assembly and disassembly. Gelsolin is one of the most potent members of the actin-severing gelsolin/villin superfamily, as it severs with nearly 100% efficiency...
. - Actin depolymerizing proteins such as ADF/cofilinCofilinADF/cofilin is a family of actin-binding proteins which disassembles actin filaments.Three highly conserved and highly identical genes belonging to this family have been described in human and mice:...
.
The actin filament network in non-muscle cells is highly dynamic. As first proposed by Dickinson & Purich (Biophysical Journal 92: 622–631), the actin filament network is arranged with the barbed-end of each filament attached to the cell's peripheral membrane by means of clamped-filament elongation motors, the above-mentioned "actoclampins", formed from a filament barbed-end and a clamping protein (formins, VASP, Mena, WASP, and N-WASP). The primary substrate for these elongation motors is Profilin-Actin-ATP complex which is directly transferred to elongating filament ends (Dickinson, Southwick & Purich, 2002). The pointed-end of each filament is oriented toward the cell's interior. In the case of lamellipodial growth, the Arp2/3 complex generates a branched network, and in filopodia a parallel array of filaments is formed.
Actin acts as a track for myosin motor motility
MyosinMyosin
Myosins comprise a family of ATP-dependent motor proteins and are best known for their role in muscle contraction and their involvement in a wide range of other eukaryotic motility processes. They are responsible for actin-based motility. The term was originally used to describe a group of similar...
motors are intracellular ATP-dependent enzymes that bind to and move along actin filaments. Various classes of myosin motors have very different behaviors, including exerting tension in the cell and transporting cargo vesicles.
A proposed model – actoclampins track filament ends
One proposed model suggests the existence of actin filament barbed-end-tracking molecular motors termed "actoclampin." The proposed actoclampins generate the propulsive forces needed for actin-based motility of lamellipodiaLamellipodia
The lamellipodium is a cytoskeletal protein actin projection on the mobile edge of the cell. It contains a quasi-two-dimensional actin mesh; the whole structure propels the cell across a substrate...
, filopodia
Filopodia
Filopodia are slender cytoplasmic projections that extend beyond the leading edge of lamellipodia in migrating cells. They contain actin filaments cross-linked into bundles by actin-binding proteins, e.g. fascin and fimbrin. Filopodia form focal adhesions with the substratum, linking it to the...
, invadipodia, dendritic spine
Dendritic spine
A dendritic spine is a small membranous protrusion from a neuron's dendrite that typically receives input from a single synapse of an axon. Dendritic spines serve as a storage site for synaptic strength and help transmit electrical signals to the neuron's cell body...
s, intracellular
Intracellular
Not to be confused with intercellular, meaning "between cells".In cell biology, molecular biology and related fields, the word intracellular means "inside the cell".It is used in contrast to extracellular...
vesicle
Vesicle
Vesicle may refer to:In cellular biology or chemistry* Vesicle , a supramolecular assembly of lipid molecules, like a cell membrane* Synaptic vesicleIn human embryology* Auditory vesicle* Optic vesiclesIn human anatomy and morphology...
s, and motile processes in endocytosis
Endocytosis
Endocytosis is a process by which cells absorb molecules by engulfing them. It is used by all cells of the body because most substances important to them are large polar molecules that cannot pass through the hydrophobic plasma or cell membrane...
, exocytosis
Exocytosis
Exocytosis , also known as 'The peni-cytosis', is the durable process by which a cell directs the contents of secretory vesicles out of the cell membrane...
, podosome formation, and phagocytosis. Actoclampin motors also propel such intracellular pathogen
Pathogen
A pathogen gignomai "I give birth to") or infectious agent — colloquially, a germ — is a microbe or microorganism such as a virus, bacterium, prion, or fungus that causes disease in its animal or plant host...
s as Listeria monocytogenes, Shigella flexneri
Shigella flexneri
Shigella flexneri is a species of Gram-negative bacteria in the genus Shigella that can cause diarrhea in humans. There are several different serogroups of Shigella; S. flexneri belongs to group B. S. flexneri infections can usually be treated with antibiotics although some strains have become...
, Vaccinia
Vaccinia
Vaccinia virus is a large, complex, enveloped virus belonging to the poxvirus family. It has a linear, double-stranded DNA genome approximately 190 kbp in length, and which encodes for approximately 250 genes. The dimensions of the virion are roughly 360 × 270 × 250 nm, with a mass of...
and Rickettsia
Rickettsia
Rickettsia is a genus of non-motile, Gram-negative, non-sporeforming, highly pleomorphic bacteria that can present as cocci , rods or thread-like . Being obligate intracellular parasites, the Rickettsia survival depends on entry, growth, and replication within the cytoplasm of eukaryotic host cells...
. When assembled under suitable conditions, these end-tracking molecular motors can also propel biomimetic particles.
The term actoclampin is derived from acto- to indicate the involvement of an actin filament, as in actomyosin, and clamp to indicate a clasping device used for strengthening flexible/moving objects and for securely fastening two or more components, followed by the suffix -in to indicate its protein origin. An actin filament end-tracking protein may thus be termed a clampin.
Dickinson and Purich (2002) recognized that prompt ATP hydrolysis
ATP hydrolysis
ATP hydrolysis is the reaction by which chemical energy that has been stored and transported in the high-energy phosphoanhydridic bonds in ATP is released, for example in the muscles, to produce work. The product is ADP and an inorganic phosphate, orthophosphate...
could explain the forces achieved during actin-based motility. They proposed a simple mechanoenzymatic sequence known as the Lock, Load & Fire Model, in which an end-tracking protein remains tightly bound ("locked" or clamped) onto the end of one sub-filament of the double-stranded actin filament. After binding to Glycyl-Prolyl-Prolyl-Prolyl-Prolyl-Prolyl-registers on tracker proteins, Profilin-ATP-actin is delivered ("loaded") to the unclamped end of the other sub-filament, whereupon ATP
Adenosine triphosphate
Adenosine-5'-triphosphate is a multifunctional nucleoside triphosphate used in cells as a coenzyme. It is often called the "molecular unit of currency" of intracellular energy transfer. ATP transports chemical energy within cells for metabolism...
within the already clamped terminal subunit of the other subfragment is hydrolyzed ("fired"), providing the energy needed to release that arm of the end-tracker, which then can bind another Profilin-ATP-actin to begin a new monomer-addition round.
Steps involved
The following steps describe one force-generating cycle of an actoclampin molecular motor:
- The polymerization cofactor profilin and the ATP·actin combine to form a profilin-ATP-actin complex that then binds to the end-tracking unit
- The cofactor and monomer are transferred to the barbed-end of an actin already clamped filament
- The tracking unit and cofactor dissociate from the adjacent protofilament, in a step that can be facilitated by ATP hydrolysis energy to modulate the affinity of the cofactor and/or the tracking unit for the filament; and this mechanoenzymatic cycle is then repeated, starting this time on the other sub-filament growth site.
When operating with the benefit of ATP hydrolysis, AC motors generate per-filament forces of 8–9 pN, which is far greater than the per-filament limit of 1–2 pN for motors operating without ATP hydrolysis (Dickinson and Purich, 2002, 2006; Dickinson, Caro and Purich, 2004). The term actoclampin is generic and applies to all actin filament end-tracking molecular motors, irrespective of whether they are driven actively by an ATP-activated mechanism or passively.
Some actoclampins (e.g., those involving Ena/VASP proteins, WASP, and N-WASP) apparently require Arp2/3-mediated filament initiation to form the actin polymerization nucleus that is then "loaded" onto the end-tracker before processive motility can commence. To generate a new filament, Arp2/3 requires a "mother" filament, monomeric ATP-actin, and an activating domain from Listeria ActA or the VCA region of N-WASP. Ther Arp2/3 complex binds to the side of the mother filament, forming a Y-shaped branch having a 70 degree angle with respect to the longitudinal
Longitudinal
Longitudinal may refer to:* Longitudinal engine, an internal combustion engine in which the crankshaft is oriented along the long axis of the vehicle, front to back...
axis of the mother filament. Then upon activation by ActA or VCA, the Arp complex is believed to undergo a major conformational change, bringing its two actin-related protein subunits near enough to each other to generate a new filament gate. Whether ATP hydrolysis may be required for nucleation and/or Y-branch release is a matter under active investigation.