

Genetic drift
Encyclopedia
Genetic drift or allelic drift is the change in the frequency of a gene variant (allele
) in a population due to random sampling
.
The alleles in the offspring are a sample of those in the parents, and chance has a role in determining whether a given individual survives and reproduces. A population's allele frequency
is the fraction of the copies of one gene that share a particular form. Genetic drift may cause gene variants to disappear completely and thereby reduce genetic variation.
The effect of genetic drift is larger for alleles present in a smaller number of copies, and smaller when an allele is present in many copies. Vigorous debates occurred over the relative importance of natural selection
versus neutral processes, including genetic drift. Ronald Fisher
held the view that genetic drift plays at the most a minor role in evolution, and this remained the dominant view for several decades. In 1968 Motoo Kimura
rekindled the debate with his neutral theory of molecular evolution
, which claims that most instances where a genetic change spreads across a population
(although not necessarily changes in phenotypes) are caused by genetic drift.
P2 takes his 20 cans, now full of paint, off to the back of his own truck. P3 awaits him with 20 empty cans. Continue ad infinitum. Eventually, a painter will walk away with 20 cans all of the same color.
It is obvious that this can happen at any generation - if, for example, P1 had by chance always chosen one of his red cans when filling P2s empties, P2 would have walked away with 20 cans of red paint - and, therefore, would not have any blue paint to give to P3, who would have to be content with all red. Slightly less obvious is that the odds of this happening become smaller if the number of cans to be filled (that is, the size of the population) is larger. With 20 cans, the odds of P1 always picking a red can are (1/2)20 - about one in a million. Slim, but with 200 cans the odds become 1 in 1.6x1060 - less probable than picking out a specific atom from amongst all the atoms in the sun.
More cans gives us a better statistical sample
, meaning that the number of red and blue cans is likely to remain much closer to what it was in the previous generation. Genetic drift is weaker in large populations - the frequency of an allele (i.e., the fraction of cans that are red, in our example) will change more slowly between generations when the population size is large. Conversely, drift is fast in small populations - with only 4 cans, it might only take a few iterations before they are all the same color. Genetic drift thus tends to eliminate variation more quickly in small populations; large populations will tend to have greater genetic diversity
.
predicts little change taking place over time when the population is large. When the reproductive population is small, however, the effects of sampling error
can alter the allele frequencies
significantly. Genetic drift is therefore considered to be a consequential mechanism of evolutionary change primarily within small, isolated populations.
This effect can be illustrated with a simplified example. Consider a very large colony of bacteria isolated in a drop of solution. The bacteria are genetically identical except for a single gene with two alleles labeled A and B. Half the bacteria have allele A and the other half have allele B. Thus both A and B have allele frequency 1/2.
A and B are neutral alleles—meaning they do not affect the bacteria's ability to survive and reproduce. This being the case, all bacteria in this colony are equally likely to survive and reproduce. The drop of solution then shrinks until it has only enough food to sustain four bacteria. All the others die without reproducing. Among the four who survive, there are sixteen possible combination
s for the A and B alleles:
(A-A-A-A), (B-A-A-A), (A-B-A-A), (B-B-A-A),
(A-A-B-A), (B-A-B-A), (A-B-B-A), (B-B-B-A),
(A-A-A-B), (B-A-A-B), (A-B-A-B), (B-B-A-B),
(A-A-B-B), (B-A-B-B), (A-B-B-B), (B-B-B-B).
If each of the combinations with the same number of A and B respectively are counted, we get the following table. The probabilities are calculated with the slightly faulty premise that the peak population size was infinite.
The probability of any one possible combination is
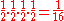
where 1/2 (the probability of the A or B allele for each surviving bacterium) is multiplied four times (the total sample size, which in this example is the total number of surviving bacteria).
As seen in the table, the total number of possible combinations to have an equal (conserved) number of A and B alleles is six, and its probability is 6/16. The total number of possible alternative combinations is ten, and the probability of unequal number of A and B alleles is 10/16.
The total number of possible combinations can be represented as binomial coefficient
s and they can be derived from Pascal's triangle
. The probability for any one of the possible combinations can be calculated with the formula

where N is the number of bacteria and k is the number of A (or B) alleles in the combination. The function '' signifies the binomial coefficient and can be expressed as "N choose k". Using the formula to calculate the probability that between them the surviving four bacteria have two A alleles and two B alleles.
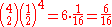
Genetic drift occurs when a population's allele frequencies change due to random events. In this example the population contracted to just four random survivors, a phenomenon known as population bottleneck
. The original colony began with an equal distribution of A and B alleles but chances are that the remaining population of four members has an unequal distribution. The probability that this surviving population will undergo drift (10/16) is higher than the probability that it will remain the same (6/16).
es or a diffusion equation describing changes in allele frequency.
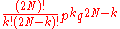
where the symbol "!" signifies the factorial
function. This expression can also be formulated using the binomial coefficient,
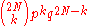
assumes overlapping generations. At each time step, one individual is chosen to reproduce and one individual is chosen to die. So in each timestep, the number of copies of a given allele can go up by one, go down by one, or can stay the same. This means that the transition matrix is tridiagonal, which means that mathematical solutions are easier for the Moran model than for the Wright-Fisher model. On the other hand, computer simulations are usually easier to perform using the Wright-Fisher model, because fewer time steps need to be calculated. In the Moran model, it takes N timesteps to get through one generation, where N is the effective population size
. In the Wright-Fisher model, it takes just one.
In practice, the Moran model and Wright-Fisher model give qualitatively similar results, but genetic drift runs twice as fast in the Moran model.
), genetic drift is a less powerful force compared to selection.
One important alternative source of stochasticity, perhaps more important than genetic drift, is genetic draft
. Genetic draft is the effect on a locus by selection on linked
loci. The mathematical properties of genetic draft are different from those of genetic drift. The direction of the random change in allele frequency is autocorrelated
across generations.
, genetic mutation
, or selection
.
Populations do not gain new alleles from the random sampling of alleles passed to the next generation, but the sampling can cause an existing allele to disappear. Because random sampling
can remove, but not replace, an allele, and because random declines or increases in allele frequency influence expected allele distributions for the next generation, genetic drift drives a population towards genetic uniformity over time. When an allele reaches a frequency of 1 (100%) it is said to be "fixed" in the population and when an allele reaches a frequency of 0 (0%) it is lost. Once an allele becomes fixed
, genetic drift comes to a halt, and the allele frequency cannot change unless a new allele is introduced in the population via mutation or gene flow
. Thus even while genetic drift is a random, directionless process, it acts to eliminate genetic variation
over time.
 Assuming genetic drift is the only evolutionary force acting on an allele, after t generations in many replicated populations, starting with allele frequencies of p and q, the variance in allele frequency across those populations is
Assuming genetic drift is the only evolutionary force acting on an allele, after t generations in many replicated populations, starting with allele frequencies of p and q, the variance in allele frequency across those populations is
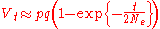
For example, if the frequency p for allele A is 75% and the frequency q for allele B is 25%, then given unlimited time the probability A will ultimately become fixed in the population is 75% and the probability that B will become fixed is 25%.
The expected number of generations for fixation to occur is proportional
to the population size, such that fixation is predicted to occur much more rapidly in smaller populations. Normally the effective population size
, which is smaller than the total population, is used to determine these probabilities. The effective population (Ne) takes into account factors such as the level of inbreeding, the stage of the lifecycle in which the population is the smallest, and the fact that some neutral genes are genetically linked to others that are under selection. The effective population size may not be the same for every gene in the same population.
One forward-looking formula used for approximating the expected time before a neutral allele becomes fixed through genetic drift, according to the Wright-Fisher model, is

where T is the number of generations, Ne is the effective population size, and p is the initial frequency for the given allele. The result is the number of generations expected
to pass before fixation occurs for a given allele in a population with given size (Ne) and allele frequency (p).
The expected time for the neutral allele to be lost through genetic drift can be calculated as
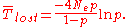
When a mutation appears only once in a population large enough for the initial frequency to be negligible, the formulas can be simplified to
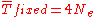
for average number of generations expected before fixation of a neutral mutation, and
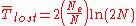
for the average number of generations expected before the loss of a neutral mutation.
nor selection
. If an allele is lost by mutation much more often than it is gained by mutation, then mutation, as well as drift, may influence the time to loss. If the allele prone to mutational loss begins as fixed in the population, and is lost by mutation at rate m per replication, then the expected time in generations until its loss in a haploid population is given by
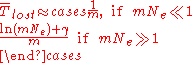
where is equal to Euler's constant. The first approximation represents the waiting time until the first mutant destined for loss, with loss then occurring relatively rapidly by genetic drift, taking time Ne<<1/m. The second approximation represents the time needed for deterministic loss by mutation accumulation. In both cases, the time to fixation is dominated by mutation via the term 1/m, and is less affected by the effective population size
is equal to Euler's constant. The first approximation represents the waiting time until the first mutant destined for loss, with loss then occurring relatively rapidly by genetic drift, taking time Ne<<1/m. The second approximation represents the time needed for deterministic loss by mutation accumulation. In both cases, the time to fixation is dominated by mutation via the term 1/m, and is less affected by the effective population size
.
s to the current environment, genetic drift has no direction and is guided only by the mathematics of chance
. As a result, drift acts upon the genotypic frequencies
within a population without regard to their phenotypic effects. In contrast, selection favors the spread of alleles whose phenotypic effects increase survival and/or reproduction of their carriers. Selection lowers the frequencies of alleles that cause unfavorable traits, and ignores those that are neutral.
In natural populations, genetic drift and natural selection
do not act in isolation; both forces are always at play. However, the degree to which alleles are affected by drift or selection varies according to population size. The magnitude of drift on allele frequencies per generation is larger in small populations. The magnitude of drift is large enough to overwhelm selection when the selection coefficient
is less than 1 divided by the effective population size
. As a result, drift affects the frequency of more alleles in small populations than in large ones.
When the allele frequency is very small, drift can also overpower selection—even in large populations. For example, while disadvantageous mutations are usually eliminated quickly in large populations, new advantageous mutations are almost as vulnerable to loss through genetic drift as are neutral mutations. Not until the allele frequency for the advantageous mutation reaches a certain threshold will genetic drift have an effect.
The mathematics of genetic drift depend on the effective population size
, but it is not clear how this is related to the actual number of individuals in a population. Genetic linkage
to other genes that are under selection can reduce the effective population size experienced by a neutral allele. With a higher recombination
rate, linkage decreases and with it this local effect on effective population size. This effect is visible in molecular data as a correlation between local recombination rate and genetic diversity, and negative correlation between gene density and diversity at noncoding sites. Stochasticity associated with linkage to other genes that are under selection is not the same as sampling error, and is sometimes known as genetic draft
in order to distinguish it from genetic drift.
is when a population contracts to a significantly smaller size over a short period of time due to some random environmental event. In a true population bottleneck, the odds for survival of any member of the population are purely random, and are not improved by any particular inherent genetic advantage. The bottleneck can result in radical changes in allele frequencies, completely independent of selection.
The impact of a population bottleneck can be sustained, even when the bottleneck is caused by a one-time event such as a natural catastrophe. After a bottleneck, inbreeding
increases. This increases the damage done by recessive deleterious mutations, in a process known as inbreeding depression
. The worst of these mutations are selected against, leading to the loss of other alleles that are genetically linked
to them, in a process of background selection. This leads to a further loss of genetic diversity. In addition, a sustained reduction in population size increases the likelihood of further allele fluctuations from drift in generations to come.
A population's genetic variation can be greatly reduced by a bottleneck, and even beneficial adaptations may be permanently eliminated.
The loss of variation leaves the surviving population vulnerable to any new selection pressures such as disease, climate change or shift in the available food source, because adapting in response to environmental changes requires sufficient genetic variation in the population for natural selection to take place.
There have been many known cases of population bottleneck in the recent past. Prior to the arrival of Europeans, North American prairies were habitat for millions of greater prairie chicken
s. In Illinois
alone, their numbers plummeted from about 100 million birds in 1900 to about 50 birds in the 1990s. The declines in population resulted from hunting and habitat destruction, but the random consequence has been a loss of most of the species' genetic diversity
. DNA analysis comparing birds from the mid century to birds in the 1990s documents a steep decline in the genetic variation in just in the latter few decades. Currently the greater prairie chicken is experiencing low reproductive success
.
Over-hunting also caused a severe population bottleneck in the northern elephant seal
in the 19th century. Their resulting decline in genetic variation can be deduced by comparing it to that of the southern elephant seal
, which were not so aggressively hunted.
A well documented example is found in the Amish
migration to Pennsylvania
in 1744. Two members of the new colony shared the recessive allele for Ellis–van Creveld syndrome. Members of the colony and their descendants tend to be religious isolates and remain relatively insular. As a result of many generations of inbreeding, Ellis-van Creveld syndrome is now much more prevalent among the Amish than in the general population.
The difference in gene frequencies between the original population and colony may also trigger the two groups to diverge
significantly over the course of many generations. As the difference, or genetic distance
, increases, the two separated populations may become distinct, both genetically
and phenetically
, although not only genetic drift but also natural selection, gene flow, and mutation contribute to this divergence. This potential for relatively rapid changes in the colony's gene frequency led most scientists to consider the founder effect (and by extension, genetic drift) a significant driving force in the evolution of new species
. Sewall Wright
was the first to attach this significance to random drift and small, newly isolated populations with his shifting balance theory
of speciation. Following after Wright, Ernst Mayr
created many persuasive models to show that the decline in genetic variation and small population size following the founder effect were critically important for new species to develop. However, there is much less support for this view today since the hypothesis has been tested repeatedly through experimental research and the results have been equivocal at best.
, Sewall Wright
. His first use of the term "drift" was in 1929, though at the time he was using it in the sense of a directed process of change, or natural selection. Random drift by means of sampling error came to be known as the "Sewall-Wright effect", though he was never entirely comfortable to see his name given to it. Wright referred to all changes in allele frequency as either "steady drift" (e.g. selection) or "random drift" (e.g. sampling error). "Drift" came to be adopted as a technical term in the stochastic
sense exclusively. Today it is usually defined still more narrowly, in terms of sampling error. Wright wrote that the "restriction of "random drift" or even "drift" to only one component, the effects of accidents of sampling, tends to lead to confusion." Sewall Wright considered the process of random genetic drift by means of sampling error equivalent to that by means of inbreeding
, but later work has shown them to be distinct.
In the early days of the modern evolutionary synthesis
, scientists were just beginning to blend the new science of population genetics with Charles Darwin
's theory of natural selection. Working within this new framework, Wright focused on the effects of inbreeding on small relatively isolated populations. He introduced the concept of an adaptive landscape in which phenomena such as cross breeding and genetic drift in small populations could push them away from adaptive peaks, which in turn allow natural selection to push them towards new adaptive peaks. Wright thought smaller populations were more suited for natural selection because "inbreeding was sufficiently intense to create new interaction systems through random drift but not intense enough to cause random nonadaptive fixation of genes."
Wright's views on the role of genetic drift in the evolutionary scheme were controversial almost from the very beginning. One of the most vociferous and influential critics was colleague Ronald Fisher
. Fisher conceded genetic drift played some role in evolution, but an insignificant one. Fisher has been accused of misunderstanding Wright's views because in his criticisms Fisher seemed to argue Wright had rejected selection almost entirely. To Fisher, viewing the process of evolution as a long, steady, adaptive progression was the only way to explain the ever increasing complexity from simpler forms. But the debates have continued between the "gradualists" and those who lean more toward the Wright model of evolution where selection and drift together play an important role.
In 1968, population geneticist Motoo Kimura
rekindled the debate with his neutral theory of molecular evolution
, which claims that most of the genetic changes are caused by genetic drift acting on neutral mutations.
The role of genetic drift by means of sampling error in evolution has been criticized by John H Gillespie and Will Provine
, who argue that selection on linked sites is a more important stochastic force.
Allele
An allele is one of two or more forms of a gene or a genetic locus . "Allel" is an abbreviation of allelomorph. Sometimes, different alleles can result in different observable phenotypic traits, such as different pigmentation...
) in a population due to random sampling
Sampling (statistics)
In statistics and survey methodology, sampling is concerned with the selection of a subset of individuals from within a population to estimate characteristics of the whole population....
.
The alleles in the offspring are a sample of those in the parents, and chance has a role in determining whether a given individual survives and reproduces. A population's allele frequency
Allele frequency
Allele frequency or Gene frequency is the proportion of all copies of a gene that is made up of a particular gene variant . In other words, it is the number of copies of a particular allele divided by the number of copies of all alleles at the genetic place in a population. It can be expressed for...
is the fraction of the copies of one gene that share a particular form. Genetic drift may cause gene variants to disappear completely and thereby reduce genetic variation.
The effect of genetic drift is larger for alleles present in a smaller number of copies, and smaller when an allele is present in many copies. Vigorous debates occurred over the relative importance of natural selection
Natural selection
Natural selection is the nonrandom process by which biologic traits become either more or less common in a population as a function of differential reproduction of their bearers. It is a key mechanism of evolution....
versus neutral processes, including genetic drift. Ronald Fisher
Ronald Fisher
Sir Ronald Aylmer Fisher FRS was an English statistician, evolutionary biologist, eugenicist and geneticist. Among other things, Fisher is well known for his contributions to statistics by creating Fisher's exact test and Fisher's equation...
held the view that genetic drift plays at the most a minor role in evolution, and this remained the dominant view for several decades. In 1968 Motoo Kimura
Motoo Kimura
was a Japanese biologist best known for introducing the neutral theory of molecular evolution in 1968. He became one of the most influential theoretical population geneticists. He is remembered in genetics for his innovative use of diffusion equations to calculate the probability of fixation of...
rekindled the debate with his neutral theory of molecular evolution
Neutral theory of molecular evolution
The neutral theory of molecular evolution states that the vast majority of evolutionary changes at the molecular level are caused by random drift of selectively neutral mutants . The theory was introduced by Motoo Kimura in the late 1960s and early 1970s...
, which claims that most instances where a genetic change spreads across a population
Fixation (population genetics)
In population genetics, fixation is the change in a gene pool from a situation where there exist at least two variants of a particular gene to a situation where only one of the alleles remains...
(although not necessarily changes in phenotypes) are caused by genetic drift.
Analogy with cans of paint
A painter P1 has 20 cans of paint, 10 red and 10 blue, in the back of his truck. Another painter (P2, representing the following generation) comes up with 20 empty cans that he wants filled. The first painter, obliging, fills an empty can from one of his own randomly selected cans, and returns his can to the back of his truck. He continues this way, filling empty cans from his (which we will pretend never run out of paint), until all 20 are filled.P2 takes his 20 cans, now full of paint, off to the back of his own truck. P3 awaits him with 20 empty cans. Continue ad infinitum. Eventually, a painter will walk away with 20 cans all of the same color.
It is obvious that this can happen at any generation - if, for example, P1 had by chance always chosen one of his red cans when filling P2s empties, P2 would have walked away with 20 cans of red paint - and, therefore, would not have any blue paint to give to P3, who would have to be content with all red. Slightly less obvious is that the odds of this happening become smaller if the number of cans to be filled (that is, the size of the population) is larger. With 20 cans, the odds of P1 always picking a red can are (1/2)20 - about one in a million. Slim, but with 200 cans the odds become 1 in 1.6x1060 - less probable than picking out a specific atom from amongst all the atoms in the sun.
More cans gives us a better statistical sample
Sample (statistics)
In statistics, a sample is a subset of a population. Typically, the population is very large, making a census or a complete enumeration of all the values in the population impractical or impossible. The sample represents a subset of manageable size...
, meaning that the number of red and blue cans is likely to remain much closer to what it was in the previous generation. Genetic drift is weaker in large populations - the frequency of an allele (i.e., the fraction of cans that are red, in our example) will change more slowly between generations when the population size is large. Conversely, drift is fast in small populations - with only 4 cans, it might only take a few iterations before they are all the same color. Genetic drift thus tends to eliminate variation more quickly in small populations; large populations will tend to have greater genetic diversity
Genetic diversity
Genetic diversity, the level of biodiversity, refers to the total number of genetic characteristics in the genetic makeup of a species. It is distinguished from genetic variability, which describes the tendency of genetic characteristics to vary....
.
Probability and allele frequency
In probability theory, the law of large numbersLaw of large numbers
In probability theory, the law of large numbers is a theorem that describes the result of performing the same experiment a large number of times...
predicts little change taking place over time when the population is large. When the reproductive population is small, however, the effects of sampling error
Sampling error
-Random sampling:In statistics, sampling error or estimation error is the error caused by observing a sample instead of the whole population. The sampling error can be found by subtracting the value of a parameter from the value of a statistic...
can alter the allele frequencies
Allele frequency
Allele frequency or Gene frequency is the proportion of all copies of a gene that is made up of a particular gene variant . In other words, it is the number of copies of a particular allele divided by the number of copies of all alleles at the genetic place in a population. It can be expressed for...
significantly. Genetic drift is therefore considered to be a consequential mechanism of evolutionary change primarily within small, isolated populations.
This effect can be illustrated with a simplified example. Consider a very large colony of bacteria isolated in a drop of solution. The bacteria are genetically identical except for a single gene with two alleles labeled A and B. Half the bacteria have allele A and the other half have allele B. Thus both A and B have allele frequency 1/2.
A and B are neutral alleles—meaning they do not affect the bacteria's ability to survive and reproduce. This being the case, all bacteria in this colony are equally likely to survive and reproduce. The drop of solution then shrinks until it has only enough food to sustain four bacteria. All the others die without reproducing. Among the four who survive, there are sixteen possible combination
Combination
In mathematics a combination is a way of selecting several things out of a larger group, where order does not matter. In smaller cases it is possible to count the number of combinations...
s for the A and B alleles:
(A-A-A-A), (B-A-A-A), (A-B-A-A), (B-B-A-A),
(A-A-B-A), (B-A-B-A), (A-B-B-A), (B-B-B-A),
(A-A-A-B), (B-A-A-B), (A-B-A-B), (B-B-A-B),
(A-A-B-B), (B-A-B-B), (A-B-B-B), (B-B-B-B).
If each of the combinations with the same number of A and B respectively are counted, we get the following table. The probabilities are calculated with the slightly faulty premise that the peak population size was infinite.
| A | B | Combinations | Probability |
| 4 | 0 | 1 | 1/16 |
| 3 | 1 | 4 | 4/16 |
| 2 | 2 | 6 | 6/16 |
| 1 | 3 | 4 | 4/16 |
| 0 | 4 | 1 | 1/16 |
The probability of any one possible combination is
where 1/2 (the probability of the A or B allele for each surviving bacterium) is multiplied four times (the total sample size, which in this example is the total number of surviving bacteria).
As seen in the table, the total number of possible combinations to have an equal (conserved) number of A and B alleles is six, and its probability is 6/16. The total number of possible alternative combinations is ten, and the probability of unequal number of A and B alleles is 10/16.
The total number of possible combinations can be represented as binomial coefficient
Binomial coefficient
In mathematics, binomial coefficients are a family of positive integers that occur as coefficients in the binomial theorem. They are indexed by two nonnegative integers; the binomial coefficient indexed by n and k is usually written \tbinom nk , and it is the coefficient of the x k term in...
s and they can be derived from Pascal's triangle
Pascal's triangle
In mathematics, Pascal's triangle is a triangular array of the binomial coefficients in a triangle. It is named after the French mathematician, Blaise Pascal...
. The probability for any one of the possible combinations can be calculated with the formula
where N is the number of bacteria and k is the number of A (or B) alleles in the combination. The function '' signifies the binomial coefficient and can be expressed as "N choose k". Using the formula to calculate the probability that between them the surviving four bacteria have two A alleles and two B alleles.
Genetic drift occurs when a population's allele frequencies change due to random events. In this example the population contracted to just four random survivors, a phenomenon known as population bottleneck
Population bottleneck
A population bottleneck is an evolutionary event in which a significant percentage of a population or species is killed or otherwise prevented from reproducing....
. The original colony began with an equal distribution of A and B alleles but chances are that the remaining population of four members has an unequal distribution. The probability that this surviving population will undergo drift (10/16) is higher than the probability that it will remain the same (6/16).
Mathematical models of genetic drift
Mathematical models of genetic drift can be solved using either branching processBranching process
In probability theory, a branching process is a Markov process that models a population in which each individual in generation n produces some random number of individuals in generation n + 1, according to a fixed probability distribution that does not vary from individual to...
es or a diffusion equation describing changes in allele frequency.
Wright-Fisher model
Consider a gene with two alleles, A or B. In diploid populations consisting of N individuals there are 2N copies of each gene. An individual can have two copies of the same allele or two different alleles. We can call the frequency of one allele p and the frequency of the other q. The Wright-Fisher model assumes that generations do not overlap. For example, annual plants have exactly one generation per year. Each copy of the gene found in the new generation is drawn independently at random from all copies of the gene in the old generation. The formula to calculate the probability of obtaining k copies of an allele that had frequency p in the last generation is thenwhere the symbol "!" signifies the factorial
Factorial
In mathematics, the factorial of a non-negative integer n, denoted by n!, is the product of all positive integers less than or equal to n...
function. This expression can also be formulated using the binomial coefficient,
Moran model
The Moran modelMoran process
A Moran process, named after Patrick Moran, is a stochastic process used in biology to describe finite populations. It can be used to model variety-increasing processes such as mutation as well as variety-reducing effects such as genetic drift and natural selection...
assumes overlapping generations. At each time step, one individual is chosen to reproduce and one individual is chosen to die. So in each timestep, the number of copies of a given allele can go up by one, go down by one, or can stay the same. This means that the transition matrix is tridiagonal, which means that mathematical solutions are easier for the Moran model than for the Wright-Fisher model. On the other hand, computer simulations are usually easier to perform using the Wright-Fisher model, because fewer time steps need to be calculated. In the Moran model, it takes N timesteps to get through one generation, where N is the effective population size
Effective population size
In population genetics, the concept of effective population size Ne was introduced by the American geneticist Sewall Wright, who wrote two landmark papers on it...
. In the Wright-Fisher model, it takes just one.
In practice, the Moran model and Wright-Fisher model give qualitatively similar results, but genetic drift runs twice as fast in the Moran model.
Other models of drift
If the variance in the number of offspring is much greater than that given by the binomial distribution assumed by the Wright-Fisher model, then given the same overall speed of genetic drift (the variance effective population sizeEffective population size
In population genetics, the concept of effective population size Ne was introduced by the American geneticist Sewall Wright, who wrote two landmark papers on it...
), genetic drift is a less powerful force compared to selection.
Random effects other than sampling error
Random changes in allele frequencies can also be caused by effects other than sampling error, for example random changes in selection pressure.One important alternative source of stochasticity, perhaps more important than genetic drift, is genetic draft
Genetic hitchhiking
Genetic hitchhiking is the process by which an allele may increase in frequency by virtue of being linked to a gene that is positively selected. Proximity on a chromosome may allow genes to be dragged along with a selective sweep experienced by an advantageous gene nearby...
. Genetic draft is the effect on a locus by selection on linked
Genetic linkage
Genetic linkage is the tendency of certain loci or alleles to be inherited together. Genetic loci that are physically close to one another on the same chromosome tend to stay together during meiosis, and are thus genetically linked.-Background:...
loci. The mathematical properties of genetic draft are different from those of genetic drift. The direction of the random change in allele frequency is autocorrelated
Autocorrelation
Autocorrelation is the cross-correlation of a signal with itself. Informally, it is the similarity between observations as a function of the time separation between them...
across generations.
Drift and fixation
The Hardy–Weinberg principle states that within sufficiently large populations, the allele frequencies remain constant from one generation to the next unless the equilibrium is disturbed by migrationGene flow
In population genetics, gene flow is the transfer of alleles of genes from one population to another.Migration into or out of a population may be responsible for a marked change in allele frequencies...
, genetic mutation
Mutation
In molecular biology and genetics, mutations are changes in a genomic sequence: the DNA sequence of a cell's genome or the DNA or RNA sequence of a virus. They can be defined as sudden and spontaneous changes in the cell. Mutations are caused by radiation, viruses, transposons and mutagenic...
, or selection
Selection
In the context of evolution, certain traits or alleles of genes segregating within a population may be subject to selection. Under selection, individuals with advantageous or "adaptive" traits tend to be more successful than their peers reproductively—meaning they contribute more offspring to the...
.
Populations do not gain new alleles from the random sampling of alleles passed to the next generation, but the sampling can cause an existing allele to disappear. Because random sampling
Random sample
In statistics, a sample is a subject chosen from a population for investigation; a random sample is one chosen by a method involving an unpredictable component...
can remove, but not replace, an allele, and because random declines or increases in allele frequency influence expected allele distributions for the next generation, genetic drift drives a population towards genetic uniformity over time. When an allele reaches a frequency of 1 (100%) it is said to be "fixed" in the population and when an allele reaches a frequency of 0 (0%) it is lost. Once an allele becomes fixed
Fixation (population genetics)
In population genetics, fixation is the change in a gene pool from a situation where there exist at least two variants of a particular gene to a situation where only one of the alleles remains...
, genetic drift comes to a halt, and the allele frequency cannot change unless a new allele is introduced in the population via mutation or gene flow
Gene flow
In population genetics, gene flow is the transfer of alleles of genes from one population to another.Migration into or out of a population may be responsible for a marked change in allele frequencies...
. Thus even while genetic drift is a random, directionless process, it acts to eliminate genetic variation
Genetic diversity
Genetic diversity, the level of biodiversity, refers to the total number of genetic characteristics in the genetic makeup of a species. It is distinguished from genetic variability, which describes the tendency of genetic characteristics to vary....
over time.
Rate of allele frequency change under drift
Time to fixation or loss
Assuming genetic drift is the only evolutionary force acting on an allele, at any given time the probability that an allele will eventually become fixed in the population is simply its frequency in the population at that time.For example, if the frequency p for allele A is 75% and the frequency q for allele B is 25%, then given unlimited time the probability A will ultimately become fixed in the population is 75% and the probability that B will become fixed is 25%.
The expected number of generations for fixation to occur is proportional
Proportionality (mathematics)
In mathematics, two variable quantities are proportional if one of them is always the product of the other and a constant quantity, called the coefficient of proportionality or proportionality constant. In other words, are proportional if the ratio \tfrac yx is constant. We also say that one...
to the population size, such that fixation is predicted to occur much more rapidly in smaller populations. Normally the effective population size
Effective population size
In population genetics, the concept of effective population size Ne was introduced by the American geneticist Sewall Wright, who wrote two landmark papers on it...
, which is smaller than the total population, is used to determine these probabilities. The effective population (Ne) takes into account factors such as the level of inbreeding, the stage of the lifecycle in which the population is the smallest, and the fact that some neutral genes are genetically linked to others that are under selection. The effective population size may not be the same for every gene in the same population.
One forward-looking formula used for approximating the expected time before a neutral allele becomes fixed through genetic drift, according to the Wright-Fisher model, is
where T is the number of generations, Ne is the effective population size, and p is the initial frequency for the given allele. The result is the number of generations expected
Expected value
In probability theory, the expected value of a random variable is the weighted average of all possible values that this random variable can take on...
to pass before fixation occurs for a given allele in a population with given size (Ne) and allele frequency (p).
The expected time for the neutral allele to be lost through genetic drift can be calculated as
When a mutation appears only once in a population large enough for the initial frequency to be negligible, the formulas can be simplified to
for average number of generations expected before fixation of a neutral mutation, and
for the average number of generations expected before the loss of a neutral mutation.
Time to loss with both drift and mutation
The formulae above apply to an allele that is already present in a population, and which is subject to neither mutationMutation
In molecular biology and genetics, mutations are changes in a genomic sequence: the DNA sequence of a cell's genome or the DNA or RNA sequence of a virus. They can be defined as sudden and spontaneous changes in the cell. Mutations are caused by radiation, viruses, transposons and mutagenic...
nor selection
Natural selection
Natural selection is the nonrandom process by which biologic traits become either more or less common in a population as a function of differential reproduction of their bearers. It is a key mechanism of evolution....
. If an allele is lost by mutation much more often than it is gained by mutation, then mutation, as well as drift, may influence the time to loss. If the allele prone to mutational loss begins as fixed in the population, and is lost by mutation at rate m per replication, then the expected time in generations until its loss in a haploid population is given by
where
is equal to Euler's constant. The first approximation represents the waiting time until the first mutant destined for loss, with loss then occurring relatively rapidly by genetic drift, taking time Ne<<1/m. The second approximation represents the time needed for deterministic loss by mutation accumulation. In both cases, the time to fixation is dominated by mutation via the term 1/m, and is less affected by the effective population sizeEffective population size
In population genetics, the concept of effective population size Ne was introduced by the American geneticist Sewall Wright, who wrote two landmark papers on it...
.
Genetic drift versus natural selection
Although both processes affect evolution, genetic drift operates randomly while natural selection functions non-randomly. While natural selection has a direction, guiding evolution towards heritable adaptationAdaptation
An adaptation in biology is a trait with a current functional role in the life history of an organism that is maintained and evolved by means of natural selection. An adaptation refers to both the current state of being adapted and to the dynamic evolutionary process that leads to the adaptation....
s to the current environment, genetic drift has no direction and is guided only by the mathematics of chance
Probability
Probability is ordinarily used to describe an attitude of mind towards some proposition of whose truth we arenot certain. The proposition of interest is usually of the form "Will a specific event occur?" The attitude of mind is of the form "How certain are we that the event will occur?" The...
. As a result, drift acts upon the genotypic frequencies
Genotype frequency
In population genetics, the genotype frequency is the frequency or proportion In population genetics, the genotype frequency is the frequency or proportion In population genetics, the genotype frequency is the frequency or proportion (i.e. 0 In population genetics, the genotype frequency is the...
within a population without regard to their phenotypic effects. In contrast, selection favors the spread of alleles whose phenotypic effects increase survival and/or reproduction of their carriers. Selection lowers the frequencies of alleles that cause unfavorable traits, and ignores those that are neutral.
In natural populations, genetic drift and natural selection
Natural selection
Natural selection is the nonrandom process by which biologic traits become either more or less common in a population as a function of differential reproduction of their bearers. It is a key mechanism of evolution....
do not act in isolation; both forces are always at play. However, the degree to which alleles are affected by drift or selection varies according to population size. The magnitude of drift on allele frequencies per generation is larger in small populations. The magnitude of drift is large enough to overwhelm selection when the selection coefficient
Selection coefficient
In population genetics, the selection coefficient is a measure of the relative fitness of a phenotype. Usually denoted by the letter s, it compares the fitness of a phenotype to another favored phenotype, and is the proportional amount that the considered phenotype is less fit as measured by...
is less than 1 divided by the effective population size
Effective population size
In population genetics, the concept of effective population size Ne was introduced by the American geneticist Sewall Wright, who wrote two landmark papers on it...
. As a result, drift affects the frequency of more alleles in small populations than in large ones.
When the allele frequency is very small, drift can also overpower selection—even in large populations. For example, while disadvantageous mutations are usually eliminated quickly in large populations, new advantageous mutations are almost as vulnerable to loss through genetic drift as are neutral mutations. Not until the allele frequency for the advantageous mutation reaches a certain threshold will genetic drift have an effect.
The mathematics of genetic drift depend on the effective population size
Effective population size
In population genetics, the concept of effective population size Ne was introduced by the American geneticist Sewall Wright, who wrote two landmark papers on it...
, but it is not clear how this is related to the actual number of individuals in a population. Genetic linkage
Genetic linkage
Genetic linkage is the tendency of certain loci or alleles to be inherited together. Genetic loci that are physically close to one another on the same chromosome tend to stay together during meiosis, and are thus genetically linked.-Background:...
to other genes that are under selection can reduce the effective population size experienced by a neutral allele. With a higher recombination
Homologous recombination
Homologous recombination is a type of genetic recombination in which nucleotide sequences are exchanged between two similar or identical molecules of DNA. It is most widely used by cells to accurately repair harmful breaks that occur on both strands of DNA, known as double-strand breaks...
rate, linkage decreases and with it this local effect on effective population size. This effect is visible in molecular data as a correlation between local recombination rate and genetic diversity, and negative correlation between gene density and diversity at noncoding sites. Stochasticity associated with linkage to other genes that are under selection is not the same as sampling error, and is sometimes known as genetic draft
Genetic hitchhiking
Genetic hitchhiking is the process by which an allele may increase in frequency by virtue of being linked to a gene that is positively selected. Proximity on a chromosome may allow genes to be dragged along with a selective sweep experienced by an advantageous gene nearby...
in order to distinguish it from genetic drift.
Population bottleneck
A population bottleneckPopulation bottleneck
A population bottleneck is an evolutionary event in which a significant percentage of a population or species is killed or otherwise prevented from reproducing....
is when a population contracts to a significantly smaller size over a short period of time due to some random environmental event. In a true population bottleneck, the odds for survival of any member of the population are purely random, and are not improved by any particular inherent genetic advantage. The bottleneck can result in radical changes in allele frequencies, completely independent of selection.
The impact of a population bottleneck can be sustained, even when the bottleneck is caused by a one-time event such as a natural catastrophe. After a bottleneck, inbreeding
Inbreeding
Inbreeding is the reproduction from the mating of two genetically related parents. Inbreeding results in increased homozygosity, which can increase the chances of offspring being affected by recessive or deleterious traits. This generally leads to a decreased fitness of a population, which is...
increases. This increases the damage done by recessive deleterious mutations, in a process known as inbreeding depression
Inbreeding depression
Inbreeding depression is the reduced fitness in a given population as a result of breeding of related individuals. It is often the result of a population bottleneck...
. The worst of these mutations are selected against, leading to the loss of other alleles that are genetically linked
Linkage disequilibrium
In population genetics, linkage disequilibrium is the non-random association of alleles at two or more loci, not necessarily on the same chromosome. It is also referred to as to as gametic phase disequilibrium , or simply gametic disequilibrium...
to them, in a process of background selection. This leads to a further loss of genetic diversity. In addition, a sustained reduction in population size increases the likelihood of further allele fluctuations from drift in generations to come.
A population's genetic variation can be greatly reduced by a bottleneck, and even beneficial adaptations may be permanently eliminated.
The loss of variation leaves the surviving population vulnerable to any new selection pressures such as disease, climate change or shift in the available food source, because adapting in response to environmental changes requires sufficient genetic variation in the population for natural selection to take place.
There have been many known cases of population bottleneck in the recent past. Prior to the arrival of Europeans, North American prairies were habitat for millions of greater prairie chicken
Greater Prairie Chicken
The Greater Prairie Chicken or Pinnated Grouse, Tympanuchus cupido, is a large bird in the grouse family. This North American species was once abundant, but has become extremely rare or extinct over much of its range due to habitat loss. There are current efforts to help this species gain the...
s. In Illinois
Illinois
Illinois is the fifth-most populous state of the United States of America, and is often noted for being a microcosm of the entire country. With Chicago in the northeast, small industrial cities and great agricultural productivity in central and northern Illinois, and natural resources like coal,...
alone, their numbers plummeted from about 100 million birds in 1900 to about 50 birds in the 1990s. The declines in population resulted from hunting and habitat destruction, but the random consequence has been a loss of most of the species' genetic diversity
Genetic diversity
Genetic diversity, the level of biodiversity, refers to the total number of genetic characteristics in the genetic makeup of a species. It is distinguished from genetic variability, which describes the tendency of genetic characteristics to vary....
. DNA analysis comparing birds from the mid century to birds in the 1990s documents a steep decline in the genetic variation in just in the latter few decades. Currently the greater prairie chicken is experiencing low reproductive success
Reproductive success
Reproductive success is defined as the passing of genes onto the next generation in a way that they too can pass those genes on. In practice, this is often a tally of the number of offspring produced by an individual. A more correct definition, which incorporates inclusive fitness, is the...
.
Over-hunting also caused a severe population bottleneck in the northern elephant seal
Northern Elephant Seal
The northern elephant seal is one of two species of elephant seal . It is a member of the family Phocidae . Elephant seals derive their name from their great size and from the male's large proboscis, which is used in making extraordinarily loud roaring noises, especially during the mating...
in the 19th century. Their resulting decline in genetic variation can be deduced by comparing it to that of the southern elephant seal
Southern Elephant Seal
The Southern Elephant Seal is one of the two extant species of elephant seal. It is both the most massive pinniped and member of the order Carnivora living today...
, which were not so aggressively hunted.
Founder effect
The founder effect is a special case of a population bottleneck, occurring when a small group in a population splinters off from the original population and forms a new one. The random sample of alleles in the just formed new colony is expected to grossly misrepresent the original population in at least some respects. It is even possible that the number of alleles for some genes in the original population is larger than the number of gene copies in the founders, making complete representation impossible. When a newly formed colony is small, its founders can strongly affect the population's genetic make-up far into the future.A well documented example is found in the Amish
Amish
The Amish , sometimes referred to as Amish Mennonites, are a group of Christian church fellowships that form a subgroup of the Mennonite churches...
migration to Pennsylvania
Pennsylvania
The Commonwealth of Pennsylvania is a U.S. state that is located in the Northeastern and Mid-Atlantic regions of the United States. The state borders Delaware and Maryland to the south, West Virginia to the southwest, Ohio to the west, New York and Ontario, Canada, to the north, and New Jersey to...
in 1744. Two members of the new colony shared the recessive allele for Ellis–van Creveld syndrome. Members of the colony and their descendants tend to be religious isolates and remain relatively insular. As a result of many generations of inbreeding, Ellis-van Creveld syndrome is now much more prevalent among the Amish than in the general population.
The difference in gene frequencies between the original population and colony may also trigger the two groups to diverge
Genetic divergence
Genetic divergence is the process in which two or more populations of an ancestral species accumulate independent genetic changes through time, often after the populations have become reproductively isolated for some period of time...
significantly over the course of many generations. As the difference, or genetic distance
Genetic distance
Genetic distance refers to the genetic divergence between species or between populations within a species. It is measured by a variety of parameters. Smaller genetic distances indicate a close genetic relationship whereas large genetic distances indicate a more distant genetic relationship...
, increases, the two separated populations may become distinct, both genetically
Genotype
The genotype is the genetic makeup of a cell, an organism, or an individual usually with reference to a specific character under consideration...
and phenetically
Phenotype
A phenotype is an organism's observable characteristics or traits: such as its morphology, development, biochemical or physiological properties, behavior, and products of behavior...
, although not only genetic drift but also natural selection, gene flow, and mutation contribute to this divergence. This potential for relatively rapid changes in the colony's gene frequency led most scientists to consider the founder effect (and by extension, genetic drift) a significant driving force in the evolution of new species
Speciation
Speciation is the evolutionary process by which new biological species arise. The biologist Orator F. Cook seems to have been the first to coin the term 'speciation' for the splitting of lineages or 'cladogenesis,' as opposed to 'anagenesis' or 'phyletic evolution' occurring within lineages...
. Sewall Wright
Sewall Wright
Sewall Green Wright was an American geneticist known for his influential work on evolutionary theory and also for his work on path analysis. With R. A. Fisher and J.B.S. Haldane, he was a founder of theoretical population genetics. He is the discoverer of the inbreeding coefficient and of...
was the first to attach this significance to random drift and small, newly isolated populations with his shifting balance theory
Shifting balance theory
The shifting balance theory of evolution is a view of evolution advocated by Sewall Wright that each of the four evolutionary forces is important to adaptive evolution...
of speciation. Following after Wright, Ernst Mayr
Ernst Mayr
Ernst Walter Mayr was one of the 20th century's leading evolutionary biologists. He was also a renowned taxonomist, tropical explorer, ornithologist, historian of science, and naturalist...
created many persuasive models to show that the decline in genetic variation and small population size following the founder effect were critically important for new species to develop. However, there is much less support for this view today since the hypothesis has been tested repeatedly through experimental research and the results have been equivocal at best.
History of the concept
The concept of genetic drift was first introduced by one of the founders in the field of population geneticsPopulation genetics
Population genetics is the study of allele frequency distribution and change under the influence of the four main evolutionary processes: natural selection, genetic drift, mutation and gene flow. It also takes into account the factors of recombination, population subdivision and population...
, Sewall Wright
Sewall Wright
Sewall Green Wright was an American geneticist known for his influential work on evolutionary theory and also for his work on path analysis. With R. A. Fisher and J.B.S. Haldane, he was a founder of theoretical population genetics. He is the discoverer of the inbreeding coefficient and of...
. His first use of the term "drift" was in 1929, though at the time he was using it in the sense of a directed process of change, or natural selection. Random drift by means of sampling error came to be known as the "Sewall-Wright effect", though he was never entirely comfortable to see his name given to it. Wright referred to all changes in allele frequency as either "steady drift" (e.g. selection) or "random drift" (e.g. sampling error). "Drift" came to be adopted as a technical term in the stochastic
Stochastic
Stochastic refers to systems whose behaviour is intrinsically non-deterministic. A stochastic process is one whose behavior is non-deterministic, in that a system's subsequent state is determined both by the process's predictable actions and by a random element. However, according to M. Kac and E...
sense exclusively. Today it is usually defined still more narrowly, in terms of sampling error. Wright wrote that the "restriction of "random drift" or even "drift" to only one component, the effects of accidents of sampling, tends to lead to confusion." Sewall Wright considered the process of random genetic drift by means of sampling error equivalent to that by means of inbreeding
Inbreeding
Inbreeding is the reproduction from the mating of two genetically related parents. Inbreeding results in increased homozygosity, which can increase the chances of offspring being affected by recessive or deleterious traits. This generally leads to a decreased fitness of a population, which is...
, but later work has shown them to be distinct.
In the early days of the modern evolutionary synthesis
Modern evolutionary synthesis
The modern evolutionary synthesis is a union of ideas from several biological specialties which provides a widely accepted account of evolution...
, scientists were just beginning to blend the new science of population genetics with Charles Darwin
Charles Darwin
Charles Robert Darwin FRS was an English naturalist. He established that all species of life have descended over time from common ancestry, and proposed the scientific theory that this branching pattern of evolution resulted from a process that he called natural selection.He published his theory...
's theory of natural selection. Working within this new framework, Wright focused on the effects of inbreeding on small relatively isolated populations. He introduced the concept of an adaptive landscape in which phenomena such as cross breeding and genetic drift in small populations could push them away from adaptive peaks, which in turn allow natural selection to push them towards new adaptive peaks. Wright thought smaller populations were more suited for natural selection because "inbreeding was sufficiently intense to create new interaction systems through random drift but not intense enough to cause random nonadaptive fixation of genes."
Wright's views on the role of genetic drift in the evolutionary scheme were controversial almost from the very beginning. One of the most vociferous and influential critics was colleague Ronald Fisher
Ronald Fisher
Sir Ronald Aylmer Fisher FRS was an English statistician, evolutionary biologist, eugenicist and geneticist. Among other things, Fisher is well known for his contributions to statistics by creating Fisher's exact test and Fisher's equation...
. Fisher conceded genetic drift played some role in evolution, but an insignificant one. Fisher has been accused of misunderstanding Wright's views because in his criticisms Fisher seemed to argue Wright had rejected selection almost entirely. To Fisher, viewing the process of evolution as a long, steady, adaptive progression was the only way to explain the ever increasing complexity from simpler forms. But the debates have continued between the "gradualists" and those who lean more toward the Wright model of evolution where selection and drift together play an important role.
In 1968, population geneticist Motoo Kimura
Motoo Kimura
was a Japanese biologist best known for introducing the neutral theory of molecular evolution in 1968. He became one of the most influential theoretical population geneticists. He is remembered in genetics for his innovative use of diffusion equations to calculate the probability of fixation of...
rekindled the debate with his neutral theory of molecular evolution
Neutral theory of molecular evolution
The neutral theory of molecular evolution states that the vast majority of evolutionary changes at the molecular level are caused by random drift of selectively neutral mutants . The theory was introduced by Motoo Kimura in the late 1960s and early 1970s...
, which claims that most of the genetic changes are caused by genetic drift acting on neutral mutations.
The role of genetic drift by means of sampling error in evolution has been criticized by John H Gillespie and Will Provine
Will Provine
William B. Provine is an American historian of science, particularly of evolutionary biology and population genetics. He is the Andrew H. and James S. Tisch Distinguished University Professor at Cornell University and is a professor in the Departments of History, Science and Technology Studies,...
, who argue that selection on linked sites is a more important stochastic force.
See also
- Allopatric speciationAllopatric speciationAllopatric speciation or geographic speciation is speciation that occurs when biological populations of the same species become isolated due to geographical changes such as mountain building or social changes such as emigration...
- Antigenic driftAntigenic driftThe immune system recognizes viruses when antigens on the surfaces of virus particles bind to immune receptors that are specific for these antigens. This is similar to a lock recognizing a key. After an infection, the body produces many more of these virus-specific receptors, which prevent...
- Gene poolGene poolIn population genetics, a gene pool is the complete set of unique alleles in a species or population.- Description :A large gene pool indicates extensive genetic diversity, which is associated with robust populations that can survive bouts of intense selection...
- Small population sizeSmall population sizeSmall populations behave differently from larger populations. They often result in population bottlenecks, which have harmful consequences for the survival of that population.-Demographic effects:...